寿命は何によって決まるのか
生物には、カゲロウのように短命のものもいれば、ヒトのように百年近く生きるものもいる。また同じヒトでも、現在は過去よりも平均寿命が長いし、現在においても、先進国の方が途上国よりも平均寿命が長い。この違いはなぜ生まれるのか。また、ヒトの寿命は今後も伸び続けるのか。寿命にまつわる謎について考えたい。

寿命とは何か
寿命(lifespan)とは、生まれてから死ぬまでの自己同一性を維持している時間的長さのことである。非生命体にも比喩的に使われることもあるが、本稿では、生命体に限定して使うことにしたい。寿命の解釈は、何の自己同一性に着目するかによって異なってくるが、生物学では、ゲノムの自己同一性に着目してこの言葉を使うことが多い。
多細胞生物の場合、細胞の生死と個体の生死を区別しなければならない。多細胞生物の体内では、体細胞が無性生殖的に増殖し、新陳代謝されるので、個々の体細胞の寿命は個体の寿命より短いのが通例である。そして、体細胞が部分的に死んでも、個体は死んだとはみなされない。体細胞分裂では、同じ遺伝子がコピーされるので、細胞ではなくて、遺伝子に注目するなら、同じ自己同一性が維持されていると解釈できるからだ。
有性生殖をする多細胞生物の場合、通常、親の遺伝子と子の遺伝子は異なり、親が死ねば、その遺伝子は消滅する。したがって、個体の寿命は、そのゲノム(DNA に含まれるすべての遺伝情報)が生成される受精の時点から消滅の時点までの期間とみなせる[1]。もとより、ゲノムの自己同一性と言っても、厳密に同じであり続けるということではない。特に非コードDNA領域[2]は変動が起きやすいが、形質発現に影響を与えないような変異は無視して差し支えない。
無性生殖の生物は、個体レベルでは死ぬことがあっても、寿命がないと言われるのは、寿命をゲノムの生成と消滅で区切って計測するなら、同じ遺伝子をコピーし続ける限り、その存在が消滅しないからだ。もとより、進化史上を振り返るまでもなく、無性生殖の生物の種が絶滅することがある。その意味でなら、無性生殖の生物にも寿命があることになる。つまり、無性生殖の生物の個体と種の関係は、有性生殖の多細胞生物における体細胞と個体の関係に等しいということである。
なぜ種によって寿命は異なるのか
寿命が外的な偶然性によって決まる無性生殖の生物はともかく、そうではない有性生殖の生物は、どのように寿命の上限を決めているのだろうか。一般的に言って、体が大きいほど、寿命が長く、また、代謝速度が低くなる傾向がある。そこから、代謝速度が高くなると寿命が短くなるという生命活動速度理論がかつて唱えられた。
この理論は、1908年にマックス・ルブナー(Max Rubner : 1854年6月2日 – 1932年4月27日)が最初に提唱し[3]、1950年代になると、代謝で発生するフリー・ラジカルが老化を惹き起こすというデナム・ハーマン(Denham Harman : 1916年2月14日 – 2014年11月25日)の説[4]によって補強されるようになった。
しかしその後、生命活動速度理論には多くの例外があることが判明した。鳥類は同じ大きさの哺乳類よりも長生きする。ダチョウのように、空を飛べなくなると、哺乳類と同じになり、同じ哺乳類でも、コウモリは他の哺乳類よりも長生きである。空を飛ぶ方が、陸上を歩くよりもより多くのエネルギーを必要とするので、呼吸と代謝はより活動的になり、それに伴って、活性酸素による酸化ストレスもより激しくなるはずである。それなのに、寿命が長くなるのはなぜなのかを生命活動速度理論はうまく説明できない。
2007年に発表された研究によると、脊椎動物の寿命を、体の大きさや系統発生の影響を補正して比較したところ、基礎代謝率が真獣類や鳥類の寿命と相関しないことが示された[5]。そこで、ジョナサン・シルバータウンは、『なぜ老いるのか、なぜ死ぬのか、進化論でわかる』で、体の大きな動物は、捕食者の餌食になりにくいから長生きするのであって、代謝速度が低いから長生きするのではないという見解を示している[6]。ハダカデバネズミが、他の齧歯類と体の大きさがほぼ同じであるにもかかわらず、寿命が非常に長いのも、地下では捕食者から比較的安全に身を守られるからだと言うのだ。
もとより、体を大きくしたり、外敵に襲われないようにしたりすることにはコストがかかるので、必ずしも良いことばかりということはない。シルバータウンの主張を、r-K選択説に基づき、繁殖戦略論的に解釈するなら、多産多死型のr戦略者は体が小さくて寿命が短く、少産少死型のK戦略者は体が大きくて寿命が長いということである。前者の種が「下手な鉄砲も数撃ちゃ当たる」という方針であるのに対して、後者の種は一発ごとに時間という資源を費やすことで命中の精度を高めようとしていると言える。
もとより、木や亀など、たくさん子供を作るにもかかわらず、成長した個体は長生きするというケースもある。これは、不確定性の高い出生時にはr戦略を採用し、幸運にも生き延びて成長した個体にはK戦略を採用するというハイブリッド型の戦略だ。K戦略者の典型であるヒトも、精子を大量に作るので、そこだけをみるなら、r戦略ということになる。進化の歴史、あるいは人類の歴史を振り返ってみても、r戦略からK戦略へという流れがある。つまり、個体発生的にも、系統発生的にも、r戦略からK戦略に重点を移すという傾向があるということである。
r-K戦略説は、ピエール=フランソワ・フェルフルスト(Pierre-François Verhulst : 1804年10月28日 – 1849年2月15日)のロジスティック方程式をもとに提唱された説で、個体数(P)の時間(t)的な変化は、以下のような微分方程式で表される。

この微分方程式の解は、以下のようになる。
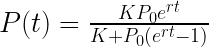
tの値が小さい時、Pはrの影響を受けて、rの指数関数のような増加関数となるが、tの値が大きくなるにつれて、PはKに近づく。一般的に言って、ロジスティック関数は、以下の図に示されるようなS字カーブを描く。最初は指数関数的に増加し、やがて環境の制約により、頭打ちになる。
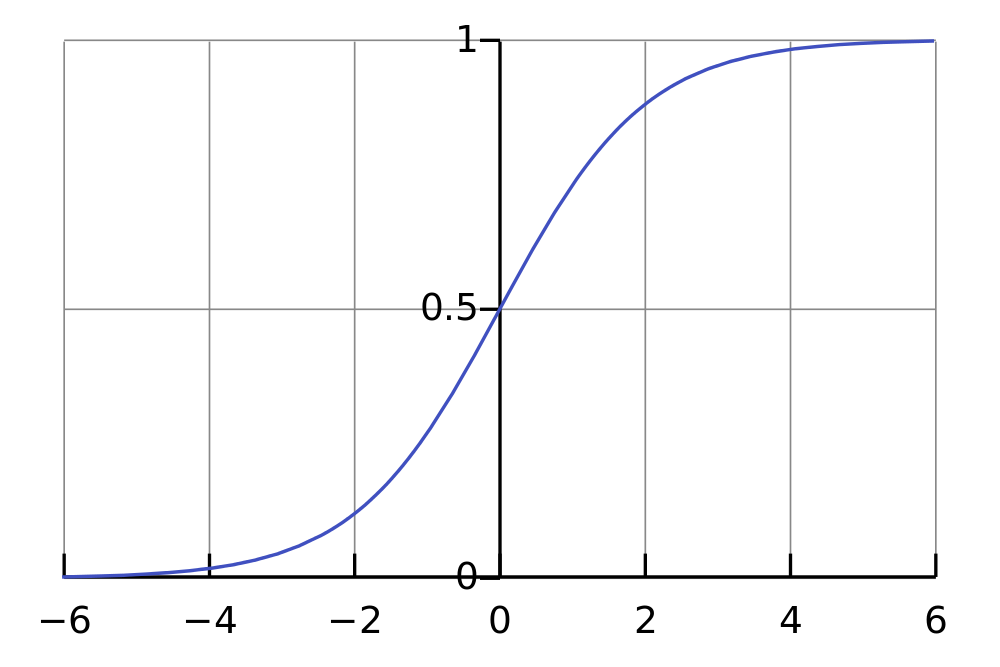
K戦略は、時間的に後になって重要になる。ただし、S字カーブはフラクタルに繰り返される。
なぜ知能が高ければ長寿になるのか
K戦略のもと、体の大きな子供を少数産むということは、一個体ごとの投資が大きくなるということであり、投資が大きくなるほどその死がもたらすサンク・コストは大きくなるので、その個体を長生きさせなければならない。それはちょうど、私たちが、安物をすぐに使い捨てにする一方で、高級品はできるだけ長く使おうとするようなものだ。もしもコストをかけても長生きしないなら、その投資は失敗ということで、自然淘汰により行われなくなる。それゆえ、自然淘汰の結果として、一個体への投資が大きい種は長寿になるということである。
体の中でも特にエネルギーを消費する臓器は脳である。したがって、脳を大きく発達させた種は、個体ごとの投資を高めていると言える。学習能力がある高等な種では、蓄積された知に資本としての価値があり、それだけに、そうした種では個体の死で生じるサンク・コストは大きくなる。霊長類が同じ大きさの哺乳類から予想されるよりも長い寿命を持つことも、その知能が高いことで説明できる。
認知能力が高くなると、それが死亡リスクに対する緩衝となり、生存確率を高めるという説を認知緩衝仮説(cognitive buffer hypothesis)と言う[8]。哺乳類においても[9]、鳥類においても[10]、脳が発達しているほど、寿命が長くなるという傾向が確認されている。カラスやオウムと言った知能の高い鳥は、同じ大きさの他の鳥よりも寿命が長い。
人類が比較的長寿であることも、その知能の高さから説明できる。また人類内部においても、知能が高ければ、長生きになるという相関が見られる[11]。2011年ごろにOECD23ヶ国を対象に行った調査によると、教育水準が高いことによる平均期待余命の延びは、25歳時点の男性で8年、女性で5年であり、65歳時点の男性で3.5年、女性で2.5年である[12]。
こう言うと、知能指数が高い、あるいは学歴の高い人は、所得が高いから長生きするのであって、知能や教育が直接寿命を延ばすことはないと反論する人もいるかもしれない。たしかに、所得と寿命には正の相関があることは事実である[13]。経済的に豊かになれば、栄養や衛生などが改善するので、寿命が延びるのは当然である。しかし、2015年に発表された米国での調査[14]によると、所得のみならず、生活習慣(喫煙や飲酒など)や心疾患リスク要因(血圧や血糖値など)の影響を取り除いて調べても、教育水準と長寿には正の相関が認められるとのことである。このことは知能や教育は、直接寿命に影響を与えるということを示している。
日本が長寿国家である原因として、治安が良く、経済的に豊かであることに加え、平均的な教育水準が高いことが挙げられる。以下の二つの図をみても、K戦略の先進国ほど寿命が長いことがわかる。


もとより、日本をはじめとする先進国は、最初からこのように平均寿命が長かったわけではない。中世における無知の状態から、知が進歩し続け、K戦略へとより大きく舵を切ることで、寿命を延ばしてきたのだ。
ヒトの寿命はなぜ延びたのか
ヒトの平均寿命、すなわち、出生時平均余命(life expectancy at birth)は、産業革命以前では40歳未満であったが、産業革命による近代化の恩恵が行き渡って以降、1年につき3か月、つまり、1時間当たり約15分のペースで延びている。2015年時点の日本人の平均寿命は83.7歳(女性は86.8歳、男性は80.5歳)で[17]、今後も同じように、1年につき3か月、つまり4年につき1歳増えていくと仮定すると、2041年には90歳を超え、2081年には100歳を超える(日本人女性だけなら、2028年に90歳を超え、2068年には100歳を超え、日本人男性だけなら、2055年に90歳を超え、2093年には100歳を超える)という計算になる。
平均寿命が線形関数的に長くなっているのは、近代化が経済を指数関数的に成長させてきたからと考えられる。というのも、以下の図が示す通り、一人当たりの購買力平価ベースの国内総生産と平均寿命の間には、対数関数的な関係が成り立つからだ[18]。
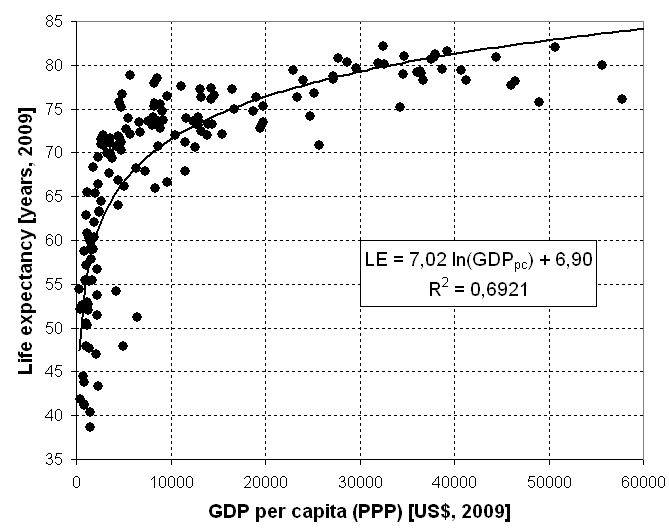
このプレストン・カーブ(Preston curve)と呼ばれる曲線は、購買力平価ベースでの収入が指数関数的に増えることで、平均寿命が線形関数的に長くなりうることを意味している。もとより、個別の値はプレストン・カーブからかなり乖離しており、経済以外の要素にも左右されることが考えられる。特に日本は、1990年代以降経済成長が停滞しているのにもかかわらず、平均寿命が延び続けており、この現象は、経済的な豊かさ以外の要因、例えば、医療技術や公衆衛生の進歩、国民のヘルス・リテラシーの向上、食品の質の改善などで説明しなければならない。
現代人が長生きになったのは、老化のペースが遅くなったからではなくて、老化のスタート時点が遅くなったからと考えられている。実際、近年「美魔女」と呼ばれる、年齢以上に若く、美しく見える女性が増えていて、それに伴って、以下の図にも見られるとおり、結婚年齢も出産年齢も上昇する傾向にあり、男性にもそれに合わせた変化が見られる。

これが寿命を延ばすことを、ゴンペルツの法則で確認しよう。ゴンペルツの法則(Gompertz law)とは、性的に成熟して以降、年齢に対する死亡率が指数関数的に増大するという法則で、この法則により、死亡率が倍になる時間、すなわち死亡率倍加時間は、一定に保たれる。死亡率倍加時間は、人では約8年、犬では約3年、実験用マウスでは約4ヶ月である[21]。以下の図は、2003年の米国における年齢ごとの死亡率をプロットしたもので、縦軸は対数目盛である。成長が止まる30歳ごろから指数関数的に死亡率が増加していることがわかる。

ゴンペルツの時代には、20歳ごろから指数関数的に増加していた。また、若者の死亡率も今よりも高かった。しかし、死亡率倍加時間は今とあまり変わらない。このことは、近代化によって平均寿命が延びたのは、死亡率倍加時間が長くなったからではなくて、老化の開始時期が遅くなり、かつ出発点の死亡率が低くなったからだと考えられる。
だが、もしもこれがヒトの平均寿命を延ばした要因だとするなら、これから先の寿命延長には限界があるということではないのか。事実、最高齢の記録は、1990年代以降更新されていないことなどから、人の平均寿命は、115歳で頭打ちになると考える研究者もいる[23]。そこで、最後に、ヒトの平均寿命を今後無限に延ばすことが可能なのかどうかについて考えてみたい。
ヒトの平均寿命は無限に延びるのか
人類史上、最も長生きをしたことが実証されているのは、ジャンヌ=ルイーズ・カルマン(Jeanne-Louise Calment : 1875年2月21日 – 1997年8月4日)で、彼女は122年と164日間生きた。このギネスにも認定されている記録は、2017年現在も破られておらず、人間の寿命の限界は120歳台にあるのではないかと考えられている。
ヒトの寿命に上限があるという定説は、生物学的には、ヘイフリック限界(Hayflick limit)に基づいている。私たちの体細胞の染色体には、テロメア (telomere) という末端部があり、テロメアは細胞分裂ごとに少しずつ短くなり、ある一定の長さにまで短くなると細胞はそれ以上分裂できなくなる[24]。ヘイフリック限界は、人の場合、50~60回で、その限界に達する時の年齢は、120~125歳とみなされている。これはカルマンの記録と合致することから、多くの生物学者から支持されている。
もっとも、ヘイフリック限界は、人間の寿命延長を阻む絶対的な制限ではない。ヒトの体内には、ヘイフリック限界に束縛されない細胞があるからだ。胚性幹細胞(ES細胞)のような多能性(Pluripotency)、すなわち、あらゆる細胞に分化する能力を持たないが、多分化能(Multipotency)、すなわち、自分が所属する特定組織における多様な細胞種へ分化可能な能力を持つ成体幹細胞(Adult stem cell 体性幹細胞とも言う)が成熟した個体内にある。例えば、造血幹細胞、神経幹細胞、皮膚幹細胞、腸管幹細胞、乳腺幹細胞など、各組織でその存在が確認されている。以下の図に示したように、成体幹細胞は、成体幹細胞と前駆細胞へと非対称分裂し、前駆細胞はヘイフリック限界の範囲内で各組織の最終分化細胞へと分裂するが、成体幹細胞は、ヘイフリック限界を超えた分裂能を示す[25]。

成体幹細胞の中でも、間葉系幹細胞は、例外的に、骨、軟骨、脂肪などの間葉系細胞のみならず、外胚葉の神経、内胚葉の肝細胞など、他の胚葉の細胞へも分化することが知られている。出澤真理らの研究チームは、この事実に着目し、間葉系組織から多能性幹細胞を同定することに成功し、この細胞をミューズ細胞(Muse cell : Multi-lineage differentiating Stress Enduring cell)と命名した[27]。ミューズ細胞は、ES細胞、iPS細胞に次ぐ「第三の多能性幹細胞[28]」として浮上している。
日本では、山中伸弥がノーベル生理学・医学賞を受賞したこともあり、iPS細胞による再生医療にばかり注目が集まりがちだが、ミューズ細胞は、iPS細胞のように分化誘導しなくても、そのまま血中に投与するだけで組織修復をもたらすという簡便性に加え、腫瘍になりにくいという安全性から、iPS細胞を上回る多能性幹細胞になるのではないかと期待されている。
STAP細胞は幻に終わった[29]が、今後も新たな多能性幹細胞の発見があるかもしれない。いずれにせよ、今後再生医学が本格的に実用化されれば、人間を不老不死化することも不可能ではなくなるだろう。神経細胞や心筋細胞は非分裂細胞なので、脳や心臓の修復しなければ、死を脳死で定義しようが、心臓死で定義しようが、どちらにせよ、人はヘイフリック限界に直面し、死ぬことになる。しかし、老化した脳や心臓を含めた全身体を多能性幹細胞で漸次修復できるようになるなら、意識の自己同一性を維持したまま、人を若返らせることが可能になる。その場合、ゴンペルツの法則は成り立たなくなるので、人の平均寿命は青天井で延び続けることになる。
不老不死を実現するもっとラディカルな方法は、マインド・アップローディング(mind uploading)あるいはマインド・トランスファー(mind transfer 精神転送)と呼ばれている方法である。人工知能が予測不可能な進歩を遂げる技術的特異点(Singularity)が近付きつつあるということで、2014年に公開された映画『トランセンデンス』のように、近年マインド・アップローディングをテーマにしたSF作品なども作られるようになった。
従来、マインド・アップローディングとして、人間の脳をスキャンし、脳の働きを人工知能でエミュレートするという、いわばソフトウェアとしての脳のリバース・エンジニアリングが提案されてきた。しかし、そうした類の「全脳エミュレーション[30]」は、たんに私たちの意識のデジタル・コピーをコンピュータ上で再現したものに過ぎず、たとえ人工知能が過去の記録をすべて引き継ぐとしても、コピーされた人の寿命が延びたとは言い難い。
寿命延長としてのマインド・アップローディングとして私が有望と考えているのは、意識の量子状態を、量子もつれ(quantum entanglement)を用いて、人間の脳と同程度以上の性能を持った量子コンピュータへと転送する方法である。量子コンピュータは、2017年にも実用化される見込みであり[31]、量子もつれを利用した情報伝達方法である量子テレポーテーション(Quantum teleportation)も、実用化に向けた実験が成功している[32]。
量子テレポーテーションは、『スタートレック』に登場するような、物体を瞬時に遠隔地へ転送するテレポーテーションとは異なり、情報だけを転送する。マインド・アップローディングに応用する場合、脳の量子状態と量子コンピュータの量子状態を、EPRペアを共有することで絡み合いの状態にして、送信側でベル測定すると、送信元の量子状態は消滅し、量子チャンネルを通じて送信先の量子コンピュータに転送される。
それではたんに意識のコピーを転送すると同時に元の意識を破壊しているだけなので、寿命延長にはなっていないと反論する人もいるかもしれない。確かに、意識が連続的、物理的に移動するわけではないから、世界で唯一無二の人格「私」の移転になっていない思う人が出てくることも無理はないが、実は、連続性がなければ自己同一性が維持できないという私たちの信念は、物理学的根拠のない思い込みである。物理的世界は、量子レベルでは非連続で、ちょうど、電光掲示板では、固定されたドットがそれぞれ点滅を繰り返しているだけなのに、文字が実体的同一性を維持したまま動いているように見えるという錯覚に陥るように、私たちは、自分たちに連続性がないのに、人格的な同一性が連続的に維持されているという幻想を抱いているだけなのである。素粒子に自己同一性がない以上、量子状態の転送で自己同一性が損なわれたと考える必要はない。
私は、第一節の冒頭で、寿命を、生まれてから死ぬまでの自己同一性を維持している時間的長さと定義した。生物学では、ゲノムの自己同一性に着目するのだが、マインド・アップローディングでは、転送先にゲノムがない以上、死んだと判断せざるをえない。しかし、私たちヒトのような意識を持った生物の場合、ゲノムの自己同一性よりも意識の自己同一性の方が重要である。もしも、マインド・アップローディングが実用化される時代が来るなら、私たちの寿命として、意識の自己同一性に着目して、寿命を再定義することになるだろう。
参照情報
- ↑私たちは、自分たちの寿命を出産のときからカウントしているが、ゲノムの自己同一性という観点から生物学的に厳密に定義するなら、私たちのゲノムが誕生した受精の時からカウントするべきだということになる。受精の時から数えても、出産の時から数えても大差はないから、確認のしやすさという実用性を重視して出産の時から数える現在の習慣に何ら問題はないが、どちらを選ぶかで大きな違いが出ることがある。今から2000年以上前の実から発芽した大賀ハスは、受精から数えるか、出芽から数えるかで、2000年以上の違いができる。
- ↑タンパク質をコード化するために使われないことから、かつてはジャンクDNA(Junk DNA ガラクタ遺伝子)と呼ばれていたが、遺伝子の発現、複製の開始、組換えが起きるホットスポットの指定、染色体凝縮、染色体分配など様々な機能があることがわかってきたため、非コードDNA領域(noncoding DNA sequences)と呼ばれるようになっている。
- ↑Max Rubner. Das Problem der Lebensdauer und seine Beziehung zu Wachstum und Ernährung. München Oldenberg. 1908. 但し、生命活動速度理論(Rate-of-living theory)という英語表記の呼称は、Raymond Pearl. The Rate Of Living. 1928. に由来する。
- ↑Harman, Denham. “Aging: a theory based on free radical and radiation chemistry." (1955): 298-300.
- ↑De Magalhães, João Pedro, Joana Costa, and George M. Church. “Analysis of the Relationship Between Metabolism, Developmental Schedules, and Longevity Using Phylogenetic Independent Contrasts." The Journals of Gerontology Series A: Biological Sciences and Medical Sciences 62.2 (2007): 149-160.
- ↑Jonathan Silvertown. The Long and the Short of It: The Science of Life Span and Aging. University of Chicago Press (2013/11/1). 8. “Live Fast, Die Young: Pace”.
- ↑Qef. “The logistic sigmoid function.” 2 July. 2008.
- ↑Sol, Daniel. “The cognitive-buffer hypothesis for the evolution of large brains." Cognitive ecology II (2009): 111-134.
- ↑C. González-Lagos, D. Sol, S. M. Reader. “Large-brained mammals live longer“. Journal of Evolutionary Biology, 23 (5), 1064-1074. 2010 May.
- ↑Sol, Daniel, et al. “Big-brained birds survive better in nature." Proceedings of the Royal Society of London B: Biological Sciences 274.1611 (2007): 763-769.
- ↑Hart, C. L., I. J. Deary, M. D. Taylor, P. L. MacKinnon, G. Davey Smith, L. J. Whalley, V. Wilson, D. J. Hole, and J. M. Starr. “The Scottish mental survey 1932 linked to the Midspan studies: a prospective investigation of childhood intelligence and future health.” Public Health 117, no. 3 (May 2003): 187–95.
- ↑Murtin, Fabrice, Johan Mackenbach, Domantas Jasilionis, and Marco Mira d’Ercole. “Inequalities in longevity by education in OECD countries.” OECD Statistics Working Papers. Paris: Organisation for Economic Co-operation and Development, January 14, 2017.
- ↑Neumayer, Eric, and Thomas Plümper. “Inequalities of Income and Inequalities of Longevity: A Cross-Country Study." American journal of public health 106.1 (2016): 160-165.
- ↑Kaplan, Robert M., et al. “Educational Attainment and Longevity: Results from the REGARDS US National Cohort Study of Blacks and Whites." Annals of epidemiology 25.5 (2015): 323-328.
- ↑Cmglee. “Comparison of male and female life expectancy at birth for countries and territories as defined in the 2011 CIA Factbook.” Licensed under CC-BY-SA.
- ↑Fobos92. “Esperanza de vida.” Licensed under CC-BY-SA.
- ↑World Health Statistics 2016: Monitoring Health for the SDGs Sustainable Development Goals (World Health Statistics Annual). World Health Organization; 1版 (2016/7/14). p.8.
- ↑Preston, S. H. “The Changing Relation between Mortality and Level of Economic Development“. Population Studies. Vol. 29, No. 2 (Jul. 1975), pp. 231-248.
- ↑Davius. “Life expectancy vs. GDP (PPP) per capita, accordig to World Bank, 2009 (sample: 162 coutries).” Licensed under CC-BY-SA. Data Sources: GDP per capita (PPP), (World Bank, 2009) Life expancy (CIA factbook, 2009).
- ↑内閣府. “晩婚化、晩産化の進行.”『平成27年版 少子化社会対策白書』第1部, 第1章, 第1節.
- ↑Finch, Caleb E. Longevity, Senescence, and the Genome. University of Chicago Press, 1994. p.23.
- ↑Uscitizenjason. “File:USGompertzCurve.” Licensed under CC-BY-SA.
- ↑Dong, Xiao, Brandon Milholland, and Jan Vijg. “Evidence for a limit to human lifespan." Nature 538.7624 (2016): 257-259.
- ↑Hayflick, L., and P. S. Moorhead. “The Serial Cultivation of Human Diploid Cell Strains.” Experimental Cell Research 25, no. 3 (December 1, 1961): 585–621.
- ↑Seruya, Mitchel, Anup Shah, Dawn Pedrotty, Tracey du Laney, Ryan Melgiri, J Andrew McKee, Henry E Young, Laura E Niklason. “Clonal Population of Adult Stem Cells: Life Span and Differentiation Potential." Cell transplantation 13.2 (2004): 93-101.
- ↑Original: “Diagram of stem cell division and differentiation.”
- ↑Kuroda, Yasumasa, et al. “Unique multipotent cells in adult human mesenchymal cell populations." Proceedings of the National Academy of Sciences 107.19 (2010): 8639-8643.
- ↑国立研究開発法人新エネルギー・産業技術総合開発機構. “新たな「ヒト多能性幹細胞(Muse細胞)」を発見 ―ES細胞、iPS細胞に次ぐ第三の多能性幹細胞―.” バイオテクノロジー・医療技術開発部. 2010年4月20日.
- ↑De Los Angeles, Alejandro, Francesco Ferrari, Yuko Fujiwara, Ronald Mathieu, Soohyun Lee, Semin Lee, Ho-Chou Tu, et al. “Failure to replicate the STAP cell phenomenon.” Nature 525, no. 7570 (September 24, 2015): E6–9.
- ↑Bostrom, Nick, and Anders Sandberg. “Whole brain emulation: a roadmap." Technical Report #2008‐3. Future of Humanity Institute, Oxford University.
- ↑Castelvecchi, D. “Quantum computers ready to leap out of the lab in 2017." Nature 541.7635 (2017): 9.
- ↑Grosshans, Frédéric. “Quantum communications: Teleportation becomes streetwise." Nature Photonics 10, 623–625 (2016).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}