ウイルス進化説は正しいか
ウイルス進化説とは、進化はウイルスの感染によって起こるという仮説のことである。「ウイルス進化説」あるいは「ウイルス進化論」は、中原英臣と佐川峻による命名であるが、この仮説は海外では二人が提唱する前からあり、それらを含めた包括的な説とするなら、今日ウイルス由来の遺伝子が哺乳類の進化をもたらしたことが実証されているので、部分的には正しいということがわかっている。

1. ウイルス進化説の起源と背景
二十世紀において支配的であった進化論のパラダイムは、ネオ・ダーウィニズム(Neo-Darwinism)と呼ばれる学説で、ダーウィンの進化論とメンデルの遺伝学を総合しているため、総合進化説(modern synthesis 現代の総合)とも呼ばれる。19世紀後半にアウグスト・ヴァイスマンが獲得形質の遺伝を否定[1]して以来、遺伝子がどのように変化して進化をもたらすのかが焦点となったが、1901年にユーゴー・ド・フリースが突然変異を発見し[2]、遺伝子突然変異で生まれた多様性が自然淘汰によって選別され、進化が起きるというネオ・ダーウィニズムが支持を集めるようになった。
1927年にハーマン・マラーが、X線で人為的に突然変異を惹き起こせることを発見し[3]、1937年には、ブレークスリーとアベリーが、化学物質(コルヒチン)によっても突然変異(倍数化)が起きることを発見した[4]。こうした放射線や化学物質といった変異原で誘発される遺伝子のコピー・ミスなどの事故的な変異は古典的な遺伝子突然変異であり、これを狭義の突然変異と呼ぶことにしよう。
その後、遺伝子は、もっと自発的に見える方法で変更されることが分かった。1941年に、バーバラ・マクリントックがトウモロコシでトランスポゾン(染色体上を移動することができる塩基配列)の転移が原因である遺伝子の変異を発見した[5]。1970年には、ハワード・マーティン・テミン[6]とデビッド・ボルティモア[7]によって逆転写酵素が見出された。これにより、遺伝情報は DNA から RNA への転写によって一方向に行われるというセントラル・ドグマが崩壊し、以下の図に示されるように、レトロウイルスも逆転写により cDNA(相補的 DNAを作り、それを宿主のDNAに組み込むことが分かった。

図中の番号に従って解説しよう。
- レトロウイルスには、プロテアーゼ(水色)、逆転写酵素(赤色)、インテグラーゼ(紺色)の3つの酵素を持つRNA鎖がある。
- 宿主細胞の受容体に吸着し、
- RNA鎖と酵素が宿主細胞に入る。
- 細胞内で、レトロウイルスRNAから逆転写酵素によりcDNAを逆転写し、
- cDNA は二本鎖DNAを形成し、核に入り、
- インテグラーゼによって宿主細胞DNAに組み込まれる。
- RNAがDNAをコピーして、転写によりmRNAを生産し、
- mRNAがリボソームでタンパク質を合成し、プロテアーゼが切断する。
- これらの断片は集められ、新しいレトロウイルスとして細胞膜から発芽する。
さらに、トランスポゾンによるコピー・アンド・ペイストであるレトロポゾンも、レトロウイルスが起源ではないかと考えられるようになった。レトロウイルスが起源と考えられるゲノムは、内在性レトロウイルス(Endogenous retrovirus)と呼ばれ、ヒト・ゲノムの 5~8% を占めるとみなされている[9]。カット・アンド・ペイストであるDNAトランスポゾンも、ゲノム内に残存したDNAウイルスが起源なのかもしれない。こうした類の遺伝子の変異は、狭義の突然変異ではないものの、「遺伝子突然変異とは、遺伝子を形成しているDNA配列の恒久的な変異である[10]」という定義に文字通り従うなら、広義の遺伝子突然変異であると言える。
レトロウイルスが逆転写によってDNAを改変することができるなら、これが進化をもたらすという考えが信憑性を持つことになる。実際、テミンとボルティモアが1970年7月出版の『ネイチャー』に逆転写酵素発見の論文を掲載したのに続いて、次の号に当たる1970年9月出版の『ネイチャー』に、生物の進化がウイルス感染による遺伝子の水平移動に大いに依存していると主張するノーマン・アンダーソンの仮説がレターとして掲載された[11]。ウイルス進化説が著名な学術誌に掲載されたのは、おそらくこれが世界で最初にちがいない。
ウイルス感染が進化を惹き起こすとアンダーソンが考えた根拠は以下の七つである。
- もしもウイルス感染が病気をもたらすだけで、宿主となる生物にとって何の有用な機能をも持たないなら、もっと感染を予防するように生物が進化しているはずだが、そうではない生物が淘汰されないということは、ウイルス感染が宿主にとって何らかの利点があることを示唆している。
- いくつかのウイルスは細胞特異性を示すものの、多くのウイルスは生物の種、あるいはしばしば門の障壁を容易に横断して感染する。
- ウイルスが宿主のDNA断片を組み込み、それを他の細胞に転移させるという事実は周知のとおりである。
- ウイルスの全てのゲノムが、生殖細胞に組み込まれ、次の世代に遺伝する可能性がある。
- 異なった種において似通った進化が起きる並行進化は、個別で起きる遺伝子の偶然的変異よりもウイルスの種を超えた感染によって容易に説明できる。
- 種を超えた遺伝コードの普遍性も、ウイルスの種を超えた感染によって容易に説明できる。。
- 適応的な進化は、小さな突然変異の積み重ねよりも、まとまったゲノムの変更によっての方が起きやすい。
最後の第7命題について補足しよう。狭義の事故的な遺伝子突然変異をキーボード入力の間違えに譬えるとするならば、ウイルス感染による遺伝子突然変異は他人が書いた文章のランダムなコピー・アンド・ペイストということになる。ランダムにするなら、無意味な文章になる可能性が高い。しかし、日本語のローマ字入力をランダムにすると、文字化けしたような無意味な文字列になるのが普通であるのに対して、コピー・アンド・ペイストの場合、コピーされる文字列が有意味であるだけに、文章全体として意味が通じる可能性は比較的高い。だからウイルス感染による遺伝子突然変異の方が、適合的な進化を惹き起こしやすいということである。
2. 日本におけるウイルス進化説
日本では、ウイルス進化説は、中原英臣と佐川峻による著作で表明された進化論仮説として知られている。以下に彼らの代表的な著作を列挙しよう。
- 1986年『ヒトはなぜ進化するのか―ダーウィンの適者生存説を覆す』
- 1987年『ヒトはなぜ人になったか―ダーウィン理論を超えたウイルス進化論』
- 1991年『進化論が変わる―ダーウィンをゆるがす分子生物学』
- 1996年『ウイルス進化論―ダーウィン進化論を超えて 』:1986年の著作の文庫版
- 1997年『生命進化の鍵はウイルスが握っていた―ここまで見えてきた進化の謎』
- 2008年『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』:1991年の著作の改訂版
当初彼らは、「今西進化論」を意識して、「ウイルス進化論」と名乗っていた。しかし、日本では、進化論の仮説は、用不用説、自然選択説、隔離説、定向進化説、総合進化説、中立進化説というように、説を付けるのが一般的になっているので、2008年の著作では、その慣習に従って「ウイルス進化説」という言葉を使っている。英語圏ではそうした慣習はなく、“Horizontal gene transfer in evolution 進化における遺伝子の水平移動”といった表現で言及される。“Viral evolution ウイルス進化”を略した“Virolution”といった造語もあるが、定着していない。本稿は日本語の著作なので、アンダーソン以来の仮説をまとめて「ウイルス進化説」と呼ぶことにしたい。
中原と佐川によると、「ウイルス進化論は1971年の今西錦司との往復書簡の中ではじめて公にした[12]」とのことである。非公開の手紙に書いたからといって「公にした」と言えるかどうかは疑問だが、仮にこれが公開であると認めたとしても、アンダーソンが論文を公開した1970年よりも後であり、どのみち、彼らにプライオリティはない。彼らはそれがよくわかっているからこそ、オリジナリティを出すために、ウイルス進化説を今西進化論と結びつけようとした。
今西進化論とは、日本の生態学者である今西錦司(1902年1月6日 – 1992年6月15日)がダーウィンの進化論に対抗して打ち出した理論で、進化は、「生存競争」、「自然淘汰」、「適者生存」ではなくて、主体的な共存である「棲み分け」によって、そして、個体単位ではなくて、種単位で、変わるべきときがきたら一斉に変わることで遂げられると主張する。もっとも自然選択(natural selection 自然淘汰)によらずして、種全体が変わるべきときがきたら一斉に変わるのはどのようなメカニズムによるのかに関して、今西進化論は何も答えない。中原と佐川は、ウイルスによる大規模感染がそれを可能にすると考えたわけである。
中原と佐川は、今西進化論の観点から、ダーウィンの進化論に対して以下の四つの疑問を投げかける。
[Q2]劣った性質をもつ個体は淘汰され、すぐれた性質をもつ個体の生き残るチャンスが大きいとは本当なのか?
[Q3]小さな突然変異と自然淘汰による個体の変化の小さな積み重なりで、新しい種の誕生というような大きな変化が、実際に起こるものなのか?
[Q4]発掘された化石の示す事実は、ダーウィンの進化論の描くシナリオとつじつまがあっているのか?[13]
しかし、こうしたダーウィニズム批判は、ダーウィンに対する誤解に基づいている。通俗的なダーウィン批判に対する反論は、「生命はいかにして進化するのか」で既に書いたことだが、重複を厭わず、これらに反論しよう。
[Q1]ここで中原と佐川が批判している突然変異とは、狭義の遺伝子突然変異である。アンダーソンの第7命題にあるとおり、ウイルスが惹き起こす突然変異の方が適応的である確率は高いが、両者の違いは程度の差に過ぎない。どちらも有害な形質を発現しうるのだから、それは自然淘汰によって除外しなければならない。それとも、彼らは、ウイルスは「進化のための器官[14]」だから、宿主が時期を見計らって、ウイルスを利用して遺伝子を変えていると思っているのだろうか。
遺伝子の水平移動は、今西進化論の「生物は変わるべきときがきたら変わる」というコンセプトを実証するひとつの具体例だろう。もっとも生物が変わるべきときがくるまでは、ウイルスはむしろあまり活勤しないだろう。しかし、一度、変わるべきときがくれば、生物はウイルスを使って遺伝子を次々と伝えることで、みんなで一緒に変わるのではないだろうか。[15]
しかし、実際のウイルスは、宿主の利益などとは無関係に、恒常的に宿主への侵入し、ランダムに遺伝子を改変することを試みている。ウイルス感染の結果起こる遺伝子の変異もランダムである以上、それが適応的な進化を帰結するためには、自然選択が必要になってくる。
[Q2]自然選択説では、足の遅いシマウマはライオンの餌食になることで淘汰され、速いシマウマは生き残る。ところが、今西は、ライオンは最初から狙いをつけた獲物めがけて一気に襲いかかるから、遅いから淘汰されるとか、速いから生き残るということはないと言う。
ライオンに目をつけられたシマウマは、単に運が悪かっただけのことなのである。これを今西は、「適者生存」ではなく「運者生存」と皮肉って、自然淘汰と適者生存による無方向な進化を批判している。[16]
むしろ運者生存の方が無方向な進化になるのではないかという揚げ足取りは措くとしても、進化の単位を個体ではなくて種に求める今西進化論が、個体レベルの偶然でダーウィンの進化論を否定するとはどういうことなのか。個体レベルなら、確かに、たまたま環境適合的な個体が死んだり、そうではない個体が生き残ったりすることはありうるが、種内にいる多数の個体に関しては、環境適合的な個体がそうでない個体よりも生き延びる場合が多いということは統計学的事実である。このように適者生存とは、環境適合的な個体の方が生き延びる確率が高いということであって、それだけで十分自然選択が働いたと言えるのである。
今西は、種内の個体間競争だけでなく、種間競争に対しても否定的であるが、個体間であれ、種間であれ、競争や闘争の代わりに協調や共生が見られるからといって、それはダーウィンの自然選択説に対する反論にはならない。日本では、“struggle for existence”は「生存競争」と訳されているが、これは「生存努力」という意味であって、協調や共生の方が競争や闘争よりも生き延びる確率を高めてくれるなら、前者を求めることは「生存努力」であり、その結果は自然選択による「適者生存」であると言うことができる。
中原と佐川は、2008年の著作で、「私は生存闘争という言葉を、ある生物が他の生物に依存するということや、個体が生きていくことだけでなく子孫をのこすのに成功すること(これはいっそう重要なことである)をふくませ、広義にまた比喩的な意味にもちいるということを、あらかじめいっておかねばならない[17]」という『種の起原』におけるダーウィンの言葉を引用している。それなら、「進化は競争の結果か協調の賜物か[18]」という対立構図でダーウィン進化論と今西進化論を対比させるべきではない。
[Q3]これは「進化は連続的な出来事か不連続な出来事か[19]」という問題でもある。ダーウィンは、小さな変異が少しずつ蓄積するという連続的な変異で多様性が生まれると考えたから、ダーウィン批判としては正しい。しかし、ダーウィニズムを改良したネオ・ダーウィニズム、すなわち総合進化説に対する批判としては適切ではない。遺伝子突然変異は形質に非連続な変化をもたらしうるからだ。中原と佐川は、「以下、ダーウィンの進化論といえば、いちいち断らなくともこの改良されたネオ・ダーウィニズムを指すことにする[20]」と前置きしながら、ダーウィニズム批判をネオ・ダーウィニズムに摩り替えるというトリックを行っている。
エルドリッジとグールドの断続平衡(punctuated equilibrium)説を援用するまでもなく、進化は均一な速度で連続的に進行するのではなく、環境の変化により比較的短期間に非連続的な進化が起こり、それ以外の環境が安定した長い期間は種は安定する。もしも突然変異が個体にランダムに起き、しかもその大半が有害なら、短期間のうちに変化した環境に種が適応できる進化はいかにして可能なのかということになる。
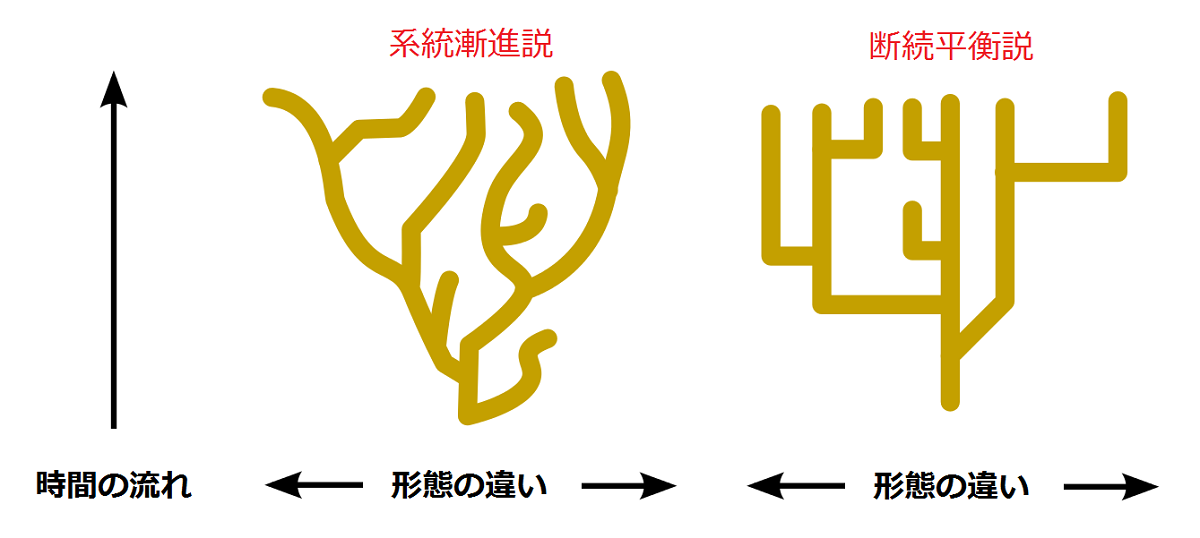
この総合進化説に対してよく投げかけられる質問は、中立進化説によって答えられる。突然変異は自然選択に対して中立もしくは「ほぼ中立[21]」であるため、淘汰されずに変異は種内で蓄積されていく。だから長期間の環境安定期に種内で多様性が生まれ、環境激変期に淘汰が起きて、変化に対応できる種内のグループが生き残り、それが新しい種を作り出す。こう考えるなら、種は変わるべきときがきたら一斉に変わるという今西の主張は、総合進化説と矛盾しないことになる。むしろ、自然淘汰を認めないなら、種が変わるべきときに一斉に変わることの説明に窮する。
中原と佐川は、ネアンデルタール人は絶滅せずに、ウイルスのおかげで、現生人類に変容した[22]と言うが、ネアンデルタール人が突如として現生人類に進化する方がよほどの奇跡でも起きない限り不可能である。実際には、ネアンデルタール人と現生人類は共通祖先から分岐し、両者の相違は自然選択にほぼ中立的だったから長い間共存できたが、最終的にはネアンデルタール人は絶滅した。現生人類は、ネアンデルタール人との間に遺伝子の交雑があったとする説[23]もあるが、だからといって、ネアンデルタール人がそのまま現生人類に進化したということにはならない。
[Q4]もしもダーウィンの連続的進化論が正しいなら、進化している途中の中間的な化石が見つかるはずだが、そのような証拠はない。しかし、ネオ・ダーウィニズムが連続的進化論ではない以上、連続的に変化する化石を証拠として挙げる必要はない。もちろん連続的ではないといっても、段階的に進化が起きる時は、中間的な化石があるはずだ。中原と佐川が、中間的な化石がないと言って好んで取り上げるのは、キリンの首が長くなる途中の化石である[24]。
キリンの首が長くなったのは、用不用説が主張するように、高い位置にある木の葉を食べようと首を伸ばしているうちに伸びたのでもなければ、自然選択説が主張するように、たまたま少し首が長かったキリンが高い位置にある木の葉を食べることができたから、自然選択により生き残ったということを何回も繰り返したのではなく、首が長くなる遺伝子を持ったウイルスに感染したことで、一挙に首が長くなったと彼らは主張する。そのようなウイルスがあるかどうかを探す前に、本当に首が中間的な長さであったキリンの祖先がいなかったのか、首が長いことが自然選択に有利に働くことはなかったのかということを考えなければならない。
キリンの先祖は、カントゥメリクスという現在のオカピに近い動物であることが分かっている。オカピは、偶蹄目キリン科オカピ属であるが、以下の写真を見てもわかるとおり、首の短い動物で、発見された当初、シマウマの一種と誤解された。

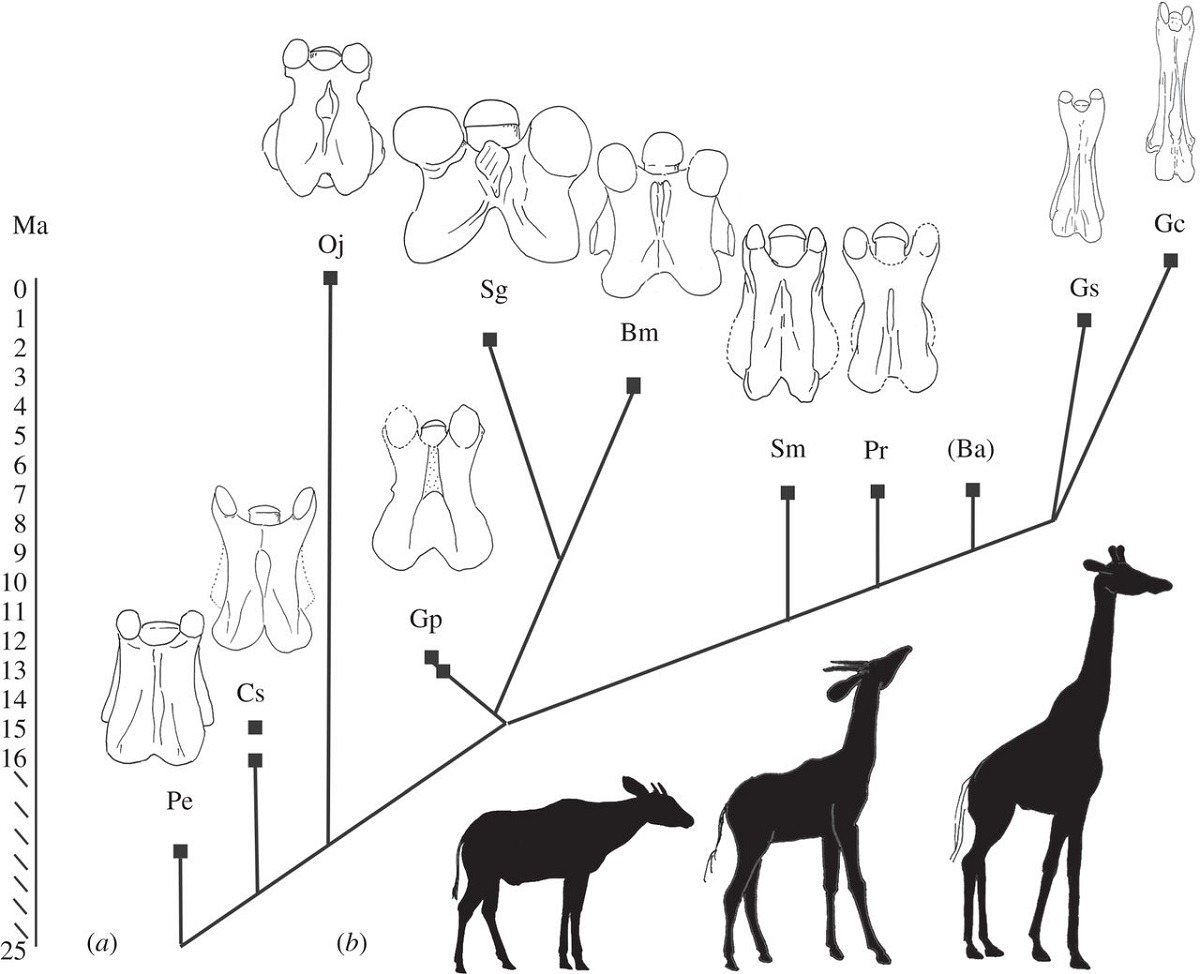
カントゥメリクス(オカピよりもやや首が長い)とキリンの中間に相当する首の長さの動物化石が見つかっている。哺乳類は、一部の例外を除いて頚椎(椎骨の最上部)が七つと決まっており、頚椎が縦長になることで、首が長くなるので、頚椎の化石の形状を見れば、その哺乳類の首の長さがわかる。以下の図は、キリン科の頚椎の進化を系統樹で描いたもので、オカピ(Oj)とキリン(Gc)以外にも、様々な形状の頚椎を持った絶滅種がいたことがわかる。絶滅種の一つ、サモテリウム(Smのシルエットを見てほしい。首の長さが、オカピとキリンの中間ぐらいであることがわかる。2015年に発表された研究によると、キリンの首は、最初の段階で脊椎の頭部側が伸び、次に脊椎の尾側が伸びたというように段階的に長くなったとのことである[26]。
キリン科の中で、最も首が短いオカピと最も首が長いキリンが生き延び、それ以外の中途半端な種が絶滅したのはなぜか。オカピが森林で暮らしているのに対して、キリンは草原で暮らしているところにヒントがある。草原では捕食動物に見つかりやすいので、捕食から免れるには、足が速くて体が大きい方が有利である。そこで、キリンは、速く走るために脚が長くなり、水を飲むために脚と同じ長さに首がなったと考えられる。これに対して、オカピは横縞模様のある保護色で森林の中で捕食者から隠れるように進化した。どっちつかずの種は、森林の中でも草原の上でも生き延びることができずに滅んだのだろう。
では、キリン以外の動物は、なぜ首が長くならなかったのか。 中原と佐川は、次のように説明している。
キリンの祖先はたまたまワンダーネットがあったので、ウイルスに感染して首が長くなっても生きていくことができた。しかしワンダーネットがない動物は、首が長くなったら生きていけなかったのである。ネコやイヌの首が長くならなかったのは、同じウイルス感染によって首が長くなったら、生きていけなかったからである。[28]
ワンダーネットがない動物は、たまたま首が長くなっても絶滅するというこの考え方は自然選択説に基づいている。自然選択による進化を否定しておきながら、自然選択で進化を説明するというのは矛盾である。
以上見てきたとおり、中原と佐川によるネオ・ダーウィニズム批判は的外れである。ウイルス進化説はむしろネオ・ダーウィニズムを補強する仮説であり、今西進化論にとって特に有利ということもない。だからウイルス進化説は、その後評価されなくなった今西進化論と運命を共にする必然性はなく、総合進化説に組み込まれる仮説として評価されるべきなのだ。
中原と佐川が、独自性を発揮しようとウイルス進化説に余計な謬見を多数くっつけたおかげで、ウイルス進化説そのものがいかがわしい理論であるかのような印象を世間に与えてしまった。もしも彼らの著作活動がなかったなら、ウイルス進化説は、たぶん「ウイルス説」とか「水平移動説」とかといった名称で、もっと素直に日本の学界で受け入れられていたことだろう。その意味で、彼らによる一連の啓蒙活動は、功罪相半ばするというよりも、むしろ罪の方が大きかったのではないかと評さざるをえない。
3. 実証されるウイルス進化説
ウイルスが宿主に感染してゲノムを変えても、それが体細胞のゲノムであるならば、ウイルス感染による遺伝子突然変異は一代限りで終わる。遺伝子突然変異が子孫に遺伝するには、ウイルスが宿主の生殖細胞のゲノムに入り込み、固定化されなければならない。また、生殖細胞から体細胞へと複製され、内在性レトロウイルスとして代々受け継がれたとしても、それがプロウイルスとして留まり、mRNAに転写されず、したがってタンパク質へと翻訳されないなら、進化を惹き起こすことはない。さらに、形質として発現しても、それが生存に対して著しく不利ならば、自然淘汰により遺伝子プールから排除されることになる。
このように、ウイルス感染自体は日常的に頻繁に起こるものの、それが進化をもたらすには越えなければならないハードルがいくつもある。だが、これらのハードルを乗り越え、実際に私たちヒトの進化に貢献した内在性レトロウイルスがあった。それはエンベロープ遺伝子である。この内在性レトロウイルスは、ヒトの場合、シンシチン(syncytin)というタンパク質をエンコードし、胎盤内の合胞体性栄養膜(syncytiotrophoblast)となることが、2000年に『ネイチャー』で発表された[29]。
哺乳類のうち、カモノハシなどの単孔類、コアラなどの有袋類以外の有胎盤類は、すべて胎盤を持つ。以下のイラストは、ヒトにおける胎盤の位置を示している。胎児がへその緒を通じて母胎とつながって円盤状の器官が胎盤である。
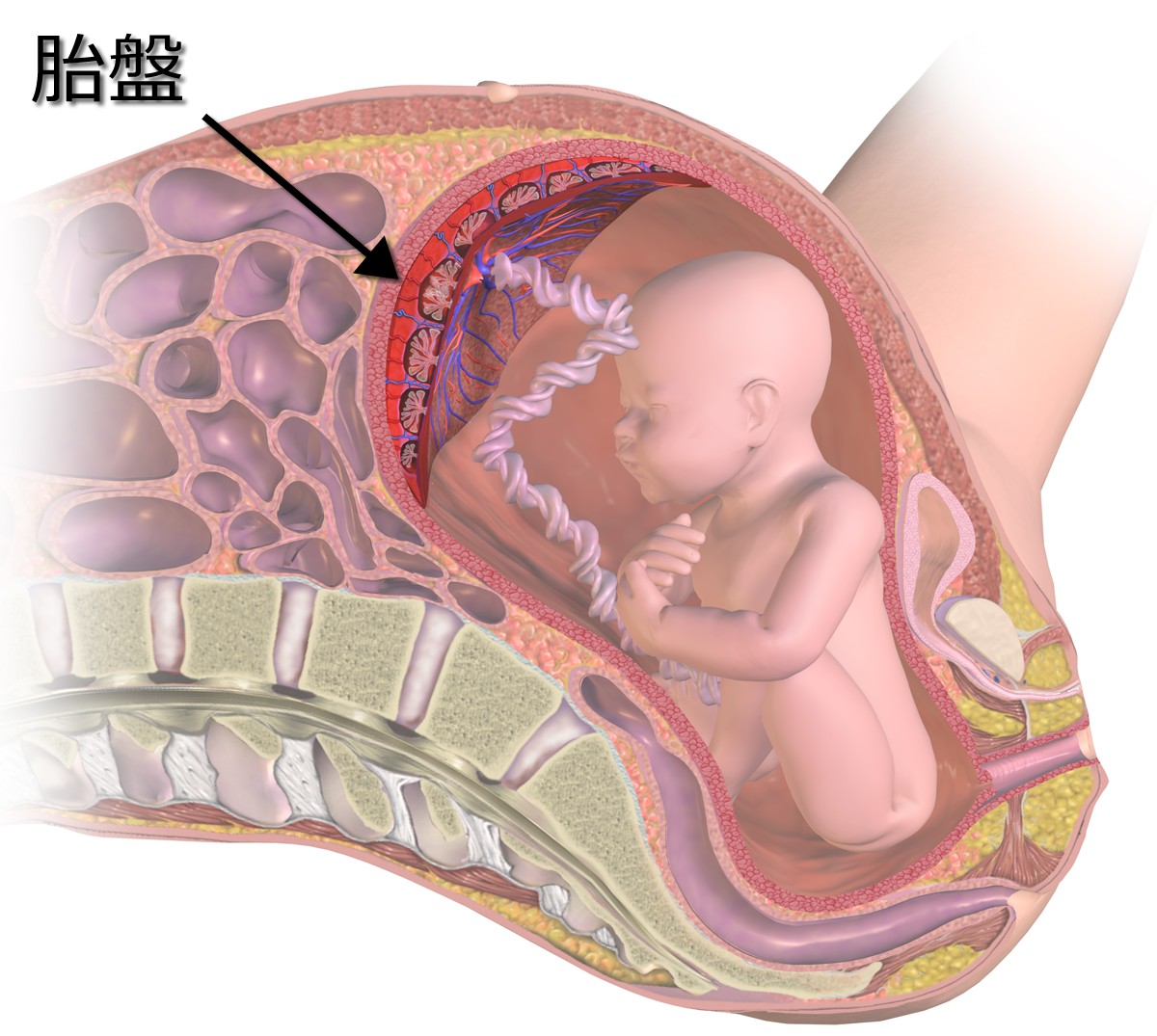
胎盤にある赤色の血管は動脈、青色の血管は静脈を表している。胎児が代謝を行うことができるように、母親は静脈から二酸化炭素と老廃物を取り除き、酸素の栄養分を動脈に供給している。その交換の場となる絨毛膜状のインターフェイスが、合胞体性栄養膜である。
合胞体(syncytium シンシチウム)とは、複数の核を含んだ細胞のことで、胎盤では、母体側の血管に接する胚由来の細胞が融合して合胞体を形成することが知られている。オスの遺伝子を分け持った胎児は母親にとって異物であり、胚と母体の間を細胞が移動することを制限する合胞体がなければ、母体の血流から免疫細胞が侵入して、胎児を攻撃してしまう。だから、有胎盤類が、有袋類とは異なり、胎盤を通して子を大きくなるまで育てるには、合胞体性栄養膜を形成する遺伝子が必要なのである。この遺伝子は、もともとレトロウイルスが宿主の細胞に侵入した際、レトロウイルスの外膜と宿主の細胞膜を融合させる働きを持っていたが、哺乳類に感染して以来、胎盤における胚栄養膜細胞の融合を活性化することに転用されている。
齧歯類は、霊長類と同じくシンシチンを作る遺伝子が合胞体性栄養膜を作る。シンシチン遺伝子をノックアウトしたマウスを使って実験してみたところ、マウスは胎盤を正常に作ることができずに流産した[31]。ここから、エンベロープ遺伝子が有胎盤類にとっていかに重要であるかがわかる。もしも私たちの祖先が、エンベロープを作るレトロウイルスに感染していなかったなら、今頃有胎盤類は存在していなかっただろう。
胎盤の形は、シンシチン遺伝子を持つヒトやマウスの場合円盤状だが、イヌやネコは帯状、ウマやブタはあちこちに散在というように、胎盤類の中でも種ごとに大きく異なる。このことは、哺乳類のレトロウイルス感染が一回ではなく複数回あって、胎盤の機能を継承(バトンタッチ)しつつも、新しいウイルス遺伝子の獲得によって、胎盤の大きさ、形、構成細胞を変化させてきたという「バトン・パス仮説[32]」が日本人研究者によって提唱されている。有胎盤類が誕生したのは、一億年ほど前のことで、生命の歴史の中では比較的最近のことだが、それ以降にもウイルスによる進化が起きていたということである。
このように、ウイルス感染によって進化が起きるというウイルス進化説は、少なくとも部分的には正しいということが実証された。ところが、2017年3月現在のウィキペディアでは、ウイルス進化説を「自然淘汰による進化を否定し、進化はウイルスの感染によって起こるという主張[33]」と定義した上で、次のように「評価」している。
本説を裏付けるに足る報告は存在せず、進化生物学の専門家からは認められた学説ではない。また、査読のある学術雑誌に投稿した論文でもないため、科学学説としても認知されていない。本説の主張は「自然選択説への誤った批判。現在までの観察、研究例の無視。非理論的な考察」によって成り立っているという批判がある。また学問的な審査を経ていないにもかかわらず、特に初学者向けの解説書などで、有力な学説であるかのように振る舞う姿勢はニセ科学に通じるとも批判される。[34]
中原と佐川の議論が「自然選択説への誤った批判。現在までの観察、研究例の無視。非理論的な考察」によって成り立っているというはそのとおりだと思うが、だからといって、ウイルス進化説をニセ科学扱いすることは正しくない。そもそも、ウイルス進化説にとって自然選択の否定は本質的な主張ではない。ちょうどダーウィンの著作に間違った内容が含まれているからといって、自然選択説の基本的主張まで否定することができないように、既存のウイルス進化説に間違った内容が含まれているからといって、ウイルス進化説の基本的主張まで否定することはできない。
中原と佐川というウイルス的な研究者のおかげで、日本の学界はウイルス進化説に対して拒絶反応を示すようになってしまった。しかし、海外の学界はウイルス進化説を取り入れ、学界のDNAともいうべき教科書までが、遺伝子の水平移動で進化が起きるというように書き換えられるようになっている。以下の図は、米国の大学で使われている進化の教科書 Evolution: Making Sense of Life の日本語訳から引用したもので、左側の図が伝統的な生物種の系統樹であるのに対して、右側は、ウイルスなどによって水平に遺伝子が伝播していることを色のついた線で表したものである。
伝統的な生物種の系統樹では、一度分岐した生物種の間で遺伝子の移動が起きないように描かれるが、それはもう古いということである。ウイルス感染以外にも、細胞質を持った生物による寄生や共生でも、遺伝子が種間で水平に移動することがあるものの、種間の遺伝子の移動でウイルスが無視できない役割を果たしたことは事実と言える。ウイルス進化説を拒絶し続けると、日本の研究者が進化論の進化に乗り遅れてしまうことになるので、ウィキペディアの「ウイルス進化説」の項目は、そろそろ書き換えた方がよいのではないか。
4. 追記:ウイルスと社会進化
2020年3月11日に世界保健機関(WHO)がパンデミックとして認定した新型コロナウイルス(SARS-CoV-2)による感染症(COVID-19)の流行は、世界を大きく変えつつある。ウイルスで命を落とす人にとって、ウイルスは悪以外の何物でもないが、本稿で述べたように、巨視的視点から見るなら、ウイルスは生命システムにとって否定的な役割だけでなく、肯定的な役割をも果たす。
一般的に言って、環境が悪化するとウイルス感染症が流行しやすくなる。これには二つの効果が感染する生命体にある。環境の悪化に伴って相対的に過剰となった個体数を減らす効果と環境の変化に適応するように遺伝的多様性を増やす効果である。細胞を乗っ取ろうとするウイルスの観点からすると、前者は失敗による効果で、後者は成功による効果である。
もっとも、2020年4月現在、新型コロナウイルス感染症による死者は世界全体で10万人未満で、今後増えるとしても、現在の医療技術の高さを考えるなら、14世紀のペスト(黒死病)や1918年のスペイン風邪のように、死者が一億人前後に到達するということはないだろう。仮に一億人が死んでも、世界の人口が77億人(2019年現在)であることを考えるなら、人口抑制効果はたかが知れる。また、人類のように、環境適応を外的な技術に依存する種にとっては、変化適応を遺伝的多様性に求める必要性は低くなる。
それでも、今回のウイルスの蔓延は、私たちに進化の機会を提供しているということができる。といっても、それは個体レベルよりもむしろ社会レベルにおいてである。「生命とは何か」でも書いたように、社会システムも一種の生命システムであり、ウイルスを契機に進化しうる。すなわち、感染爆発防止のため外出が規制される中、人間が物理的に動かなくても、情報だけを動かすことで成り立つ社会が形成されつつあるということである。
生命システムも社会システムも本来保守的で、存立の危機にでも立たされない限り、抜本的な変革はできない。ビジネス界ではしばしば「ピンチはチャンス」と言われるが、これは進化一般に言えることである。情報技術の進歩により、テレワーク(遠隔医療や遠隔教育なども含む)が技術的に可能になったが、なかなか普及しなかった。新型コロナウイルス感染症のおかげで、人々はしぶしぶテレワークを始めるようになったのだ。
新型コロナウイルス発祥の地である中国では、感染爆発のおかげで、大気汚染が激減した。ウイルスで奪われる命よりも大気汚染の低減で救われる命の方が多いのではないかという皮肉な見方もある。2019年に世界は地球温暖化で大騒ぎしたが、今回のパンデミックのおかげで、二酸化炭素の排出量は大幅に減ると予想されている。中国だけでも、25%減少したという試算がある(2020年2月中旬での評価)。
パンデミックが終われば、元に戻るという悲観的な見方もあるが、もしもテレワークが定着すれば、人類社会の環境負荷は以前よりも小さくなるだろう。情報を人間ごと動かすよりも、ネットを通じて電子的に情報だけを動かす方が、経済的コストがかからないだけでなく、環境負荷も小さくて済むからだ。テレワークを定着させることができるなら、「禍を転じて福と為す」ことができるということである。
日本では、残念ながら、そうした動きはまだあまり出ていない。グーグルが、スマートフォンの位置情報データを使って計測したモビリティ変化の報告書によると、職場へ来る動きは、2020年3月29日現在、米国では普段より38%減っているのに対し、日本は9%減るにとどまっている。3月末時点での感染の深刻さが日米で違うことも考慮しなければならないが、日本でテレワークが進まないのには、日本独特の原因もある。ハンコ文化である。テレワーク中にもかかわらず、契約書に印鑑を押すというたったそれだけのために出社するというケースもあるようだ。
オンライン上で電子契約を行うサービスもあるが、トラブルが起きた際に電子契約の内容が本当に有効なものとして扱われるかどうかを疑問視する人が多く、なかなか普及しない。改竄を心配するなら、いつ、誰が作り、誰に宛てて交付したものかをクラウド上で分散型台帳技術を用いて記録すればよいのだが、契約の当事者が新しい技術を信用しないなら、書類に印鑑を押すという古いスタイルの契約慣行が続くことになる。
テレワークだけでなく、キャッシュレス決済も感染予防のために進めなければいけない。こちらも日本は遅れているが、一般の民間企業には市場原理が働くから、まだ期待が持てる。問題は医療や教育といった行政が深く介入している規制産業の分野だ。日本の医療は対面診療が原則だが、患者が直接病院や診療所に行ったことで院内感染に巻き込まれる事例が報告されている。そこで、厚生労働省は、感染が終息するまでの時限的な措置として、初診からオンライン診療を認める方針を打ち出した。これに対して、日本医師会の横倉義武会長は、「オンラインでの初診は視覚と聴覚だけで不安だ[…]もし誤診などが出たりしたときに誰が責任を取るのか。当然、規制緩和を推進した人に取ってもらうことになるだろう」といういつもの恫喝セリフで規制緩和を牽制している。
誤診なら、従来の診断でも日常茶飯事のように起きているが、それで医者が責任を取ったという話は聞かない。従来型の診断をテレビ電話でするだけなら、オンライン診療で誤診が増えるだろうが、医療における現在のデジタル・トランスフォーメーション(DX)はもっと先を行っている。すなわち、IoTやウェアラブル端末を通して集められる患者のバイタルデータや関連ビッグデータを解析し、人工知能が高い精度で発症リスクを予測したり、疾患を診断したり、治療法の選択をアドバイスしたりできるようになっている。
こうした人を介さない医療サービスは、たんに感染リスクを低下させるだけでなく、24時間アクセス可能であることから、アクセスを制限していた従来の医療サービスと比べ、早期発見、早期治療が可能になる。これを認めないことで起きるであろう症状の悪化に対して、日本医師会や厚生労働省は責任を取らなくてよいのだろうか。もちろん、実現にはプライバシーなどの問題もあるが、プライバシーよりも健康の方を重視する人も多いであろうから、そうした人たちのために選択的に導入することは可能であろう。残念ながら、今のところ、規制緩和が時限的な措置であることからもわかるとおり、厚労省は、オンライン診療を本格化させるつもりはないようだ。
ピンチをチャンスに変え、DXを進めなければならないもう一つの規制産業は、教育である。安倍総理は、2020年2月27日に、全国全ての小学校、中学校、高等学校、特別支援学校に対して、3月2日から春休みまで臨時休業を行うよう要請した。総理が、4月7日に一部の地域に対して緊急事態宣言を発令したことから、5月6日まで休校期間が延長される地域もある。休校措置によって、子供たちから学習の機会が奪われるという問題と保護者が仕事を休んで子供の面倒を見なければならないという二つの問題が起きた。
休校措置は海外でも取られているが、ネット先進国では、学校を閉鎖しても、授業はオンラインで続けるというところが多い。日本は、遠隔授業の取り組みが遅れていたので、大半の学校は今回の事態に対処できていない。もしもネットを通じて教師あるいは人工知能が生徒の学習を監視することができるなら、休校措置に伴って発生する二つの問題の解決につながったであろう。文科省は、オンライン授業を標準授業時数に認める措置を検討しているようだが、学校の当事者には、ピンチをチャンスに変え、遠隔授業を本格化させようとする意欲はあまりないようだ。あくまでも一時的なピンチだから、すぐにまた元に戻ることができると考えているのだろう。
だが、今回のピンチを一過性の災害とみなすことはできない。登校が難しくなる事態が今後も頻発することが予想されるからだ。2018年夏の猛暑の時は、学校で熱中症が起きることが問題になった。それで教室にエアコンが設置されるようになったが、通学途中で熱中症になるリスクがあるのだから、対策として不十分である。猛暑の時、冷房の効いた自宅に留まって、遠隔授業を受けるのが最も安全な方法である。冷房がない貧困世帯には、個別に助成を行えばよい。高額な学校用エアコンを設置するよりも安くつく。
遠隔授業を行うためには、各生徒にタブレット端末を配布しなければならないので、高くつくと思う人もいるかもしれない。各生徒に一台のセルラー・タブレットを配布した渋谷区モデルでは、1人あたり27万8000円というコストの高さが話題なった。しかし、遠隔授業をするために、タブレット端末を配布する必要はない。ほとんどの家庭にはネット接続可能な端末が既にあるのだから、それをそのまま活用すればよい。現在のネットサービスはクラウド化されているので、端末の種類の違いは重要ではない。端末がない貧困家庭には、個別に助成を行えばよい。システムも、オープンソースのソフトウェアを使えば、費用を圧縮できる。もっとも、公教育は公共事業なので、費用がかかるほど儲かる人の意見が通りやすい。結局のところ、デジタル・トランスフォーメーションによって、劇的に教育のコスト・パフォーマンスを高めるために最もよい方法は、公教育の廃止であるが、既得権益を失う人が多いので、政治的には難しい。
日本は、ネットの利用に関しては、世界的に見て遅れている。『DIGITAL 2019』によると、2019年における1日のインターネット利用時間の世界平均が6時間42分であるのに対して、日本はそれが3時間45分しかない。モバイルに限定すると、世界平均が3時間14分であるのに対して、日本は1時間25分しかない。ネット普及率と回線速度は世界の平均を超えているのに、あまり積極的に活用されていないことがわかる。ネットインフラは立派だけれども、宝の持ち腐れになっているということだ。
日本経済の成長が30年以上にわたって停滞している理由の一つとして、情報革命の遅れを挙げることができる。今回のパンデミックを契機にデジタル・トランスフォーメーションを進める国と、パンデミック以前に戻ることしか考えていない守旧派の国とでは今後大きな差が付くだろう。そして、日本は、今のところ後者になりそうな見込みだ。ウイルスによって惹き起こされる人類社会の進化から取り残されると、日本は途上国になってしまうだろう。但し、途上国といっても、発展途上国という意味ではなくて、没落途上国という意味でだ。
5. 参照情報
- ↑“901 young were produced by five generations of artificially mutilated parents, and yet there was not a single example of a rudimentary tail or of any other abnormality in this organ.” Weismann, August. “On the Supposed Transmission of Mutilations” in Essays Upon Heredity and Kindred Biological Problems. p.432.
- ↑De Vries, Hugo. Die Mutationstheorie. Versuche und Beobachtungen über die Entstehung von Arten im Pflanzenreich. (1904): 789-793.
- ↑Muller, H. J. “Artificial Transmutation of the Gene.” Science 66, no. 1699 (July 22, 1927): 84–87. doi:10.1126/science.66.1699.84.
- ↑Blakeslee, Albert F., and Amos G. Avery. “Methods of inducing doubling of chromosomes in plants by treatment with colchicine." Journal of Heredity 28.12 (1937): 393-411.
- ↑McClintock, Barbara. “The stability of broken ends of chromosomes in Zea mays." Genetics 26.2 (1941): 234-282.
- ↑Temin, Howard M., and Satoshi Mizutani. “Viral RNA-dependent DNA Polymerase: RNA-dependent DNA Polymerase in Virions of Rous Sarcoma Virus.” Nature 226, no. 5252 (27 June 1970): 1211–3. doi:10.1038/2261211a0. PMID 4316301.
- ↑“Two independent groups of investigators have found evidence of an enzyme in virions of RNA tumour viruses which synthesizes DNA from an RNA template. This discovery, if upheld, will have important implications not only for carcinogenesis by RNA viruses but also for the general understanding of genetic transcription: apparently the classical process of information transfer from DNA to RNA can be inverted.” Baltimore, David. “Viral RNA-dependent DNA Polymerase: RNA-dependent DNA Polymerase in Virions of RNA Tumour Viruses." Nature. 226, no. 5252 (27 June 1970): 1209–11. doi:10.1038/2261209a0. PMID 4316300.
- ↑“A retrovirus has a membrane that contains glycoproteins, which are able to bind to a receptor protein on a host cell. Within the cell there are two strands of RNA that have three enzymes, protease, reverse transcriptase, and integrase (1). The first step of replication is the binding of the glycoprotein to the receptor protein (2). Once these have been bound the cell membrane degrades and becomes part of the host cell, and the RNA strands and enzymes go into the cell (3). Within the cell, reverse transcriptase creates a complementary strand of DNA from the retrovirus RNA and the RNA is degraded, this strand of DNA is known as cDNA (4). The cDNA is then replicated, and the two strands form a weak bond and go into the nucleus (5). Once in the nucleus, the DNA is integrated into the host cells DNA with the help of integrase (6). This cell can either stay dormant, or RNA may be synthesized from the DNA and used to create the proteins for a new retrovirus (7). Ribosome units are used to transcribe the mRNA of the virus into the amino acid sequences which can be made into proteins in the Rough Endoplasmic Reticulum. This step will also make viral enzymes and capsid proteins (8). Viral RNA will be made in the nucleus. These pieces are then gathered together and are pinched off of the cell membrane as a new retrovirus (9).” Mrdavis21. “Life Cycle of a Retrovirus.” Licensed under CC-BY-SA.
- ↑“Endogenous retroviruses (ERVs) represent the proviral phase of exogenous retroviruses that have integrated into the germ line of their host. They typically consist of an internal region with three genes (gag, pol, and env) plus two flanking, noncoding LTRs, which are identical at the time of integration. Human ERVs (HERVs) comprise ≈5–8% of the human genome, with 98,000 elements and fragments…” Belshaw, Robert, Vini Pereira, Aris Katzourakis, Gillian Talbot, Jan Pačes, Austin Burt, and Michael Tristem. “Long-term reinfection of the human genome by endogenous retroviruses." Proceedings of the National Academy of Sciences of the United States of America 101, no. 14 (2004): 4894-4899.
- ↑“A gene mutation is a permanent alteration in the DNA sequence that makes up a gene.” “What is a gene mutation and how do mutations occur? – Genetics Home Reference.” U.S. National Library of Medicine.
- ↑Anderson, Norman G. “Evolutionary Significance of Virus Infection.” Nature 227, no. 5265 (September 26, 1970): 1346–47. doi:10.1038/2271346a0.
- ↑中原英臣, 佐川峻.『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』. 講談社 (2008/4/22). p.241.
- ↑中原英臣, 佐川峻.『ヒトはなぜ人になったか―ダーウィン理論を超えたウイルス進化論』. 経済界 (1987/07). p.102-103.『ウイルス進化論―ダーウィン進化論を超えて 』. 早川書房 (1996/7/31). p.131-132. 『生命進化の鍵はウイルスが握っていた―ここまで見えてきた進化の謎』. 河出書房新社 (1997/02). p.62. にも同じようなダーウィン批判がある。
- ↑中原英臣, 佐川峻.『ウイルス進化論―ダーウィン進化論を超えて 』. 早川書房 (1996/7/31). p.23.
- ↑中原英臣, 佐川峻.『ウイルス進化論―ダーウィン進化論を超えて 』. 早川書房 (1996/7/31). p.97.
- ↑中原英臣, 佐川峻.『生命進化の鍵はウイルスが握っていた―ここまで見えてきた進化の謎』. 河出書房新社 (1997/02). p.65.
- ↑中原英臣, 佐川峻.『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』. 講談社 (2008/4/22). p.160.
- ↑中原英臣, 佐川峻.『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』. 講談社 (2008/4/22). p.158.
- ↑中原英臣, 佐川峻.『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』. 講談社 (2008/4/22). p.174.
- ↑中原英臣, 佐川峻.『ヒトはなぜ人になったか―ダーウィン理論を超えたウイルス進化論』. 経済界 (1987/07). p.99-100.
- ↑Ohta, Tomoko. “The Nearly Neutral Theory of Molecular Evolution." Annual Review of Ecology and Systematics 23.1 (1992): 263-286.
- ↑中原英臣, 佐川峻.『ウイルス進化論―ダーウィン進化論を超えて 』. 早川書房 (1996/7/31). p.157.
- ↑Wall, Jeffrey D., Kirk E. Lohmueller, and Vincent Plagnol. “Detecting ancient admixture and estimating demographic parameters in multiple human populations." Molecular biology and evolution 26, no. 8 (2009): 1823-1827.
- ↑中原英臣, 佐川峻.『生命進化の鍵はウイルスが握っていた―ここまで見えてきた進化の謎』. 河出書房新社 (1997/02). p.21.
- ↑Raul654. “An Okapi taken at Disney’s Animal Kingdom on January 16, 2005.” Licensed under CC-BY-SA.
- ↑“We find that cervical elongation is anisometric and unexpectedly precedes Giraffidae. Within the family, cranial vertebral elongation is the first lengthening stage observed followed by caudal vertebral elongation, which accounts for the extremely long neck of the giraffe.” Danowitz, Melinda, Aleksandr Vasilyev, Victoria Kortlandt, and Nikos Solounias. “Fossil evidence and stages of elongation of the Giraffa camelopardalis neck.” Open Science 2, no. 10 (October 1, 2015): 150393. doi:10.1098/rsos.150393.
- ↑Danowitz, Melinda, Aleksandr Vasilyev, Victoria Kortlandt, and Nikos Solounias. “Fossil evidence and stages of elongation of the Giraffa camelopardalis neck.” Open Science 2, no. 10 (October 1, 2015): 150393. doi:10.1098/rsos.150393. Figure 5. Cladogram with geological age and dorsal view of C3 vertebrae of taxa evaluated.
- ↑中原英臣, 佐川峻.『新・進化論が変わる―ゲノム時代にダーウィン進化論は生き残るか』. 講談社 (2008/4/22). p.216-217.
- ↑“Many mammalian viruses have acquired genes from their hosts during their evolution. The rationale for these acquisitions is usually quite clear: the captured genes are subverted to provide a selective advantage to the virus. Here we describe the opposite situation, where a viral gene has been sequestered to serve an important function in the physiology of a mammalian host. This gene, encoding a protein that we have called syncytin, is the envelope gene of a recently identified human endogenous defective retrovirus, HERV-W. We find that the major sites of syncytin expression are placental syncytiotrophoblasts, multinucleated cells that originate from fetal trophoblasts. We show that expression of recombinant syncytin in a wide variety of cell types induces the formation of giant syncytia, and that fusion of a human trophoblastic cell line expressing endogenous syncytin can be inhibited by an anti-syncytin antiserum. Our data indicate that syncytin may mediate placental cytotrophoblast fusion in vivo, and thus may be important in human placental morphogenesis.” Mi, Sha, et al. “Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis." Nature 403.6771 (2000): 785-789.
- ↑BruceBlaus. “Placenta.” Licensed under CC-BY-SA. Blausen Medical. 日本語の書き込みは私による。
- ↑“By generating knockout mice, we show here that homozygous syncytin-A null mouse embryos die in utero between 11.5 and 13.5 days of gestation. Refined cellular and subcellular analyses of the syncytin-A-deficient placentae disclose specific disruption of the architecture of the syncytiotrophoblast-containing labyrinth, with the trophoblast cells failing to fuse into an interhemal syncytial layer. Lack of syncytin-A-mediated trophoblast cell fusion is associated with cell overexpansion at the expense of fetal blood vessel spaces and with apoptosis, adding to the observed maternofetal interface structural defects to provoke decreased vascularization, inhibition of placental transport, and fetal growth retardation, ultimately resulting in death of the embryo. These results demonstrate that syncytin-A is essential for trophoblast cell differentiation and syncytiotrophoblast morphogenesis during placenta development, and they provide evidence that genes captured from ancestral retroviruses have been pivotal in the acquisition of new, important functions in mammalian evolution.” Dupressoir, Anne, et al. “Syncytin-A knockout mice demonstrate the critical role in placentation of a fusogenic, endogenous retrovirus-derived, envelope gene." Proceedings of the National Academy of Sciences 106.29 (2009): 12127-12132.
- ↑Imakawa, Kazuhiko, So Nakagawa, and Takayuki Miyazawa. “Baton pass hypothesis: successive incorporation of unconserved endogenous retroviral genes for placentation during mammalian evolution.” Genes to Cells 20, no. 10 (October 1, 2015): 771–88. doi:10.1111/gtc.12278.
- ↑Wikipedia. “ウイルス進化説.” 平成28年10月24日 (月) 20:49. 古いバージョン(平成21年6月5日)では、たんに「進化は伝染病である」という主張を紹介するだけだった。「自然淘汰による進化を否定し」の部分は、たんにウイルス進化説をニセ科学扱いするために付け加えたのではないか。ウイルス進化説は、読んで字の如く、ウイルスが進化をもたらすという説であり、それ以外の非本質的な主張を定義に含めるべきではない。
- ↑Wikipedia. “ウイルス進化説.” 平成28年10月24日 (月) 20:49.「自然選択説への誤った批判。現在までの観察、研究例の無視。非理論的な考察」は、中原英臣、佐川峻、富塚孝.「ウイルス進化論に対する反論にこたえる」に対する、同誌編集部による付記(Networks in Evolutionary Biology 第5号. 1987年. 52-53頁)より引用。「特に初学者向けの解説書などで、有力な学説であるかのように振る舞う姿勢はニセ科学に通じるとも批判される」には、[要出典]のタグがつけられているが、引用文からは削除した。
- ↑カール・ジンマー, ダグラス・エムレン. trans. 更科 功, 石川 牧子, 国友 良樹.『カラー図解 進化の教科書 第1巻 進化の歴史』. 講談社 (2016/11/16). ブルーバックス. p.144.
{kind=link}
{kind=link}
{kind=link}