高木由臣の有性生殖論
ゾウリムシの研究者、高木由臣は、2014年に出版した著書『有性生殖論』で、ゾウリムシのオートガミーを根拠に、有性生殖の機能を遺伝子の多様化に求める通説を否定し、有性生殖を、無性的一倍体化と無性的二倍体化を起源とする、蓄積した突然変異の有用性を検証する仕組みとする新説を提示している。高木の新説は正しいのかどうかを考えよう。

有性生殖の本来の機能は何か
高木は、無性生殖とは「性と無関係に遺伝的に均質な同類を増やす方式」であり、有性生殖とは「性が関与して遺伝的に多様な同類を生む方式」というのが「われわれの常識的な理解」であると言いつつ、次のように述べ、有性生殖の役割に対するそうした従来の見解に異議を申し立てる。
有性生殖は「性が関与して多様性を生む方式」であるというのは、簡潔にして的を射た説明であると思われるかもしれない。ところが本書では「性が関与しない有性生殖」や「多様性を生まない有性生殖」を取り上げながら、有性生殖の本質について考え、いかにして有性生殖が出現し、進化してきたかを探ろうとしている。[1]
高木が「性が関与しない有性生殖」や「多様性を生まない有性生殖」で念頭に置いているのは、彼の研究対象であるゾウリムシのオートガミー(自家生殖)やセルフィング(自系接合)である。セルフィングは、化学薬品を加えて人為的に誘導しなければ起きない接合なので、進化論的な意味はない。しかし、オートガミーは自然状態で起きるがゆえに、おそらく歴史の古い生殖法と考えられ、それゆえ無視することはできない。
オートガミーとは、ゾウリムシなどの原生動物が行っている生殖法の一つで、同じ細胞内の同じ核に由来した二つの核が融合する現象である。ゾウリムシの中には、パラメキウム・カウダトゥム(Paramecium caudatum 狭義のゾウリムシ)やミドリゾウリムシ(Paramecium bursaria)のように、接合だけする種もあるが、ヒメゾウリムシ(Paramecium aurelia)やヨツヒメゾウリムシ(Paramecium tetraurelia)のように、接合とオートガミーの両方する種もある。以下、ヨツヒメゾウリムシを例に取ることにしよう。
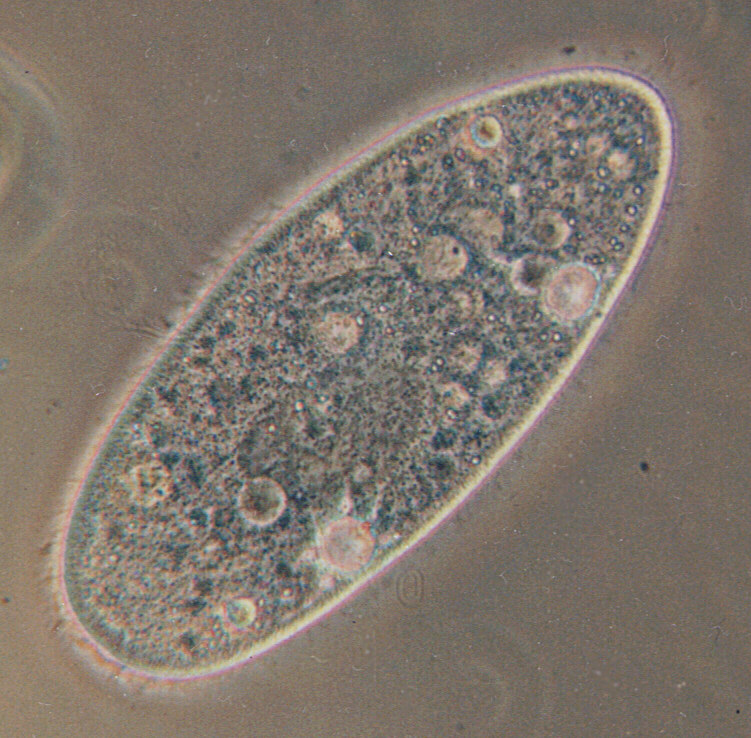
ヨツヒメゾウリムシは、餌が豊富な時は、通常の無性生殖(細胞分裂)を行って数を増やし、餌が不足して飢餓に陥ると、接合やオートガミーを行う。高木は、接合もオートガミーも、減数分裂して世代交代(若返り)を果たすから有性生殖と考えているが、接合は異なる細胞間で行う遺伝子交換であるからよいとしても、自分だけで行うオートガミーはどうなのか。それを確認するべく、以下の図でオートガミーを説明しよう。

ゾウリムシは、大核と小核という二種類の細胞核を持ち、そのうち、小核は生殖を司り、大核はそれ以外の機能を司る。図の左側に描かれているように、オートガミーにおいては、大核は消失し、二つある小核は八つに減数分裂した後、一つだけ残った一倍体核がさらに有糸分裂し、融合し、核分化により、大核一つと小核二つを持った、元と同じゾウリムシになる。図の右側に描かれているように、途中で組み換えが起きても、遺伝子は aBc/aBc のままで、変化しない。
もっとも、遺伝子に変化をもたらさないのは、親が A/A といったホモ接合型である場合で、A/a といったヘテロ接合型の場合、子では、優性遺伝子のホモ接合と劣性遺伝子のホモ接合型が、したがってそれぞれの表現型が、[A]:[a]=1:1 の比率で現れる。ヘテロ接合型では表現されなかった劣勢の形質が現れることで、その形質の環境適応性が試される。突然変異で生じる遺伝子は劣性であることが多く、高木はオートガミーを「突然変異の検証システム」と特徴付けている。
オートガミーは一見遺伝的多様化とは無縁の、有性生殖の利点が生かされていない価値の低い有性生殖であるかのような印象を与えたかもしれないが、突然変異の検証システムという観点から見ると、実に優れた有性生殖法であることか分かる。[4]
二倍体のゾウリムシが、餌が豊富な時、通常の無性生殖(細胞分裂)を行って数を増やし、餌が不足して飢餓に陥ると、オートガミーを行う理由も、突然変異の検証システムという観点から理解できる。ちょうど企業が、景気が良い時には積極的に設備投資し、人材を雇用して事業を拡大するのに対して、不況になると、採算性の悪い事業を選んで撤退を決めたり、成績の悪い人材を選別して、リストラしたりするように、ゾウリムシは、餌が豊富な時は、劣性遺伝子が蓄積しても、それには目をつぶり、ひたすら数を増やすが、餌が不足すると、オートガミーによって劣性形質を焙り出し、環境適応的でない表現型を淘汰するという守りの姿勢に入るというわけだ。
オートガミーは有性生殖か
ゾウリムシのオートガミーは典型的な有性生殖でもなければ、典型的な無性生殖でもない。では、どちらに分類するべきなのか。この問いに答えるには、有性生殖の本質は何であるのかを考え、有性生殖を定義しなければならない。
高木によれば、有性生殖には、性の分化、遺伝的多様化(ゲノムの多様化)、減数分裂と受精、世代交代(若返り)といった側面があり、これまで、最初の二つが有性生殖の要件として重視されてきた。その結果、この二つの要件を満たすバクテリアの接合が最古の有性生殖とみなされてきたのだが、高木は、バクテリアの接合に有性生殖の起源を求めることに反対する。
このことは、有性生殖のエッセンスは「性の分化による遺伝的多様化」ではなく、「減数分裂と受精による世代交代(若返り)」であることを強力に支持する。[5]
高木は、バクテリアには寿命がないことをも問題にする。『寿命論』という本の中で、高木は寿命を「受精に始まる自己同一性の継続期間[6]」と定義している。私も「生まれてから死ぬまでの自己同一性を維持している時間的長さ」と定義したので、この定義に異議はない。生物の場合、「自己同一性」とはゲノムの自己同一性と解釈するべきで、その場合、無性生殖であっても、接合でゲノムが変更される以上、寿命がないとは言えない。ところが、高木は、無性生殖には寿命がないことを前提に、バクテリアの接合は有性生殖ではないと言う。
バクテリアが無限に無性生殖を継続できる「寿命をもたない生物」であることは間違いない。もちろん餌があればという条件や、事故による死はここで言う意味での寿命とはみなさないという前提での話である。もし「バクテリアは寿命をもたない生物である」ことを認めるなら、「寿命をもたない生物は有性生殖をしない」のだから、「バクテリアは有性生殖をしない」「バクテリアの接合は有性生殖ではない」ことになる。もちろん「有性生殖をする生物は寿命をもつ」という命題が真理であるという前提での話である。[7]
バクテリアが接合によってゲノムを変えるといっても、それはプログラムされた変更ではないから、プログラムされた寿命を持つとは言えないということなのだろう。これに対して、ゾウリムシの無性生殖的な分裂には制限があり、ヨツヒメゾウリムシは約300回分裂すると、それ以上分裂できなくなって寿命に達する。ところが、オートガミーを行うと、分裂限界がリセットされ、また分裂を始めるようになる。それゆえ、オートガミーには、世代交代(若返り)という効果があり、その点でもオートガミーは有性生殖とみなすべきだというのだ。
以下の表は、有性生殖の4つの特徴に関する高木による整理である[8]。表1における水平遺伝とは、トランスポゾン、ウイルス感染、寄生、共生のように、上の世代から下の世代へと垂直に遺伝するのではない遺伝子の移転のことである。
| 有性生殖 | オートガミー | 細菌の接合 | 水平遺伝 | |
| 性の分化 | 〇 | × | 〇 | × |
| 遺伝的多様化 | 〇 | × | 〇 | 〇 |
| 世代交代(若返り) | 〇 | 〇 | × | × |
| 減数分裂と受精 | 〇 | 〇 | × | × |
高木は、減数分裂と受精による世代交代(若返り)を重視して有性生殖を定義し、オートガミーは有性生殖だと主張するのだが、有性生殖の起源を探るには、相同染色体の対合が欠かせない減数分裂は複雑すぎるとして、その本質を二倍体の一倍体化に求める。
私は、「減数分裂」は「二倍体の一倍体化」であり、「受精」は「一倍体の二倍体化」ではないかという認識から、減数分裂と受精という現象が見られる以前に、減数分裂と受精とは全く違ったやり方での「一倍体化と二倍体化」が行われ、それが有性生殖の原初的なプロセスであった時代があったのではないかと想定したのである。[9]
この想定に基づいて、高木は、減数分裂ではない一倍体化と受精ではない二倍体化との無性的な移行の繰り返しが有性生殖の原初的な姿であったとする「原初有性生殖仮説」なるものを打ち出す。
1倍体の無性生殖サイクルは、ゲノムの安全対策として、複製の重複により2倍体の無性生殖サイクルに移行し、2倍体の無性生殖サイクルは、蓄積した突然変異の有用性を検証するため、分配の重複により1倍体の無性生殖過程へ移行する。[10]
一倍体の単細胞生物が細胞分裂するという最も原始的な無性生殖においても、分裂に先立って遺伝子を複製するのであるから、「無性的二倍体化と無性的一倍体化」を行っていることになり、無性生殖と有性生殖の区別がつかなくなるようにも思えるのだが、おそらく高木は、二倍体になっている時間が長くなるような中間的状態を想像しているのだろう。
以下の図は、高木が考える典型的な無性生殖から典型的な有性生殖(異系交配型への段階的進化のプロセスである。ゾウリムシでは、二番目から四番目までの方の生殖形態が見られ、高木は単純な形態からより複雑な形態の順番に並べたようだ。二番目のオートガミー型と三番目の同系交配型の順番は逆でもありうるとしつつも、有性生殖は、突然変異を素早く検証する仕組みから突然変異を貯めこんで潜在的な遺伝子型の多様性を高める方向に向かって進化したと高木は考える。

自系接合とはセルフィングのことで、遺伝的に均質な同性のゾウリムシに化学薬品を加えると接合を人為的に誘導できることから、異系接合するゾウリムシにも潜在的に自系接合の能力があり、同系交配やオートガミーから異系交配へと進化した証拠と高木は考えている。
最古の有性生殖は何だったのか
以上が、高木の有性生殖論の要旨であるが、彼の議論の根本的な問題は、有性生殖の起源という進化論的なテーマを扱っているのにもかかわらず、その仮説が古生物学的に実証されていないところにある。高木は、ゾウリムシの生殖を最古の有性生殖と考えているようだが、それよりももっと古い有性生殖がある。それも、高木が取り上げたバクテリアの接合のような、有性生殖とみなしてよいかどうか意見が分かれるようなものではなくて、誰が見ても有性生殖と考えるような事例が、ゾウリムシの出現以前にある。それは、原始的な原生生物である紅藻(こうそう red algae/Rhodophyta)が行っている有性生殖である。

以下の図に示されている通り、紅藻では、減数分裂によって生じた単相の四分胞子から雌雄の配偶体が発達し、雄の配偶体が作る精子が雌の配偶体にある造果器内の卵細胞と受精し、果胞子体という複相の世代を形成し、果胞子体が放出する果胞子が、減数分裂により四分胞子体になることで生活史のサイクルを閉じる。
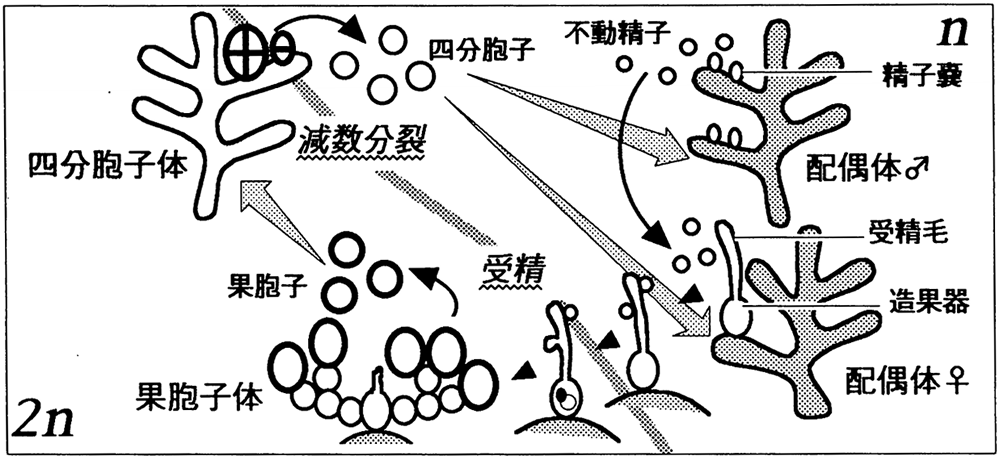
紅藻の生殖は、性の分化、遺伝的多様化(ゲノムの多様化)、減数分裂と受精、世代交代(若返り)という高木が挙げた有性生殖の四つの特徴をすべて兼ね備えており、これが有性生殖だということに異議はあるまい。
2000年に発表された論文[14]によると、カナダの北極圏にあるサマセット島で、現在のウシケノリ(Bangia)とほぼ同じ紅藻が性分化した配偶子を形成していたことを示す化石記録が、中原生代と新原生代の境界線(Mesoproterozoic/Neoproterozoic boundary)付近の地層で見つかり、これにより、12億年前に有性生殖が始まっていたことが確認された。ウシケノリには、受精した造果器自体が直接分割して、多数個の果胞子を生ずるという特徴があるが、それ以外の点では、真正紅藻類と大きくは異ならない。これは、現在知られている最も古い有性生殖の化石記録である。『有性生殖論』は、2014年に出版された本であるにもかかわらず、これに触れていない。
これに対して、ゾウリムシ属の最古の化石は、2億3000万~2億2000万年前のものと考えられている[15]。繊毛虫に対象を広げても、最古の化石は5億8千万年前である[16]。化石が発見されていなくても存在した可能性はあるが、ゾウリムシなどの原生動物は、10億年前の新原生代(Neoproterozoic)になって出てきた生物で、原始的な原生生物である紅藻類よりも前に遡ることはないのであるから、ゾウリムシのオートガミーに有性生殖の起源を求めるのはおかしい。
動物と植物は違うと思うかもしれないが、原生生物では動物と植物との間の区別は未分化で、動物だけを切り離して考える合理的根拠はない。ちなみに、動物による有性生殖の痕跡を残した最古の化石証拠は、2005年に南オーストラリア奥地の海底で発見された[17]。その化石は、約5億6500万年前の無脊椎動物、フニジア・ドロシア(Funisia dorothea)が、幼生着底していたことを窺わせる。動物に限定しても、ゾウリムシの有性生殖が最古とは言えない。
高木は、遺伝的に均質な同性のゾウリムシに化学薬品を加えると接合を人為的に誘導できることから、異系接合を行うゾウリムシにも潜在的にセルフィング(自系接合)の能力があり、異系交配はそこから進化したと考えていることは既に述べた。セルフィングは原生動物だけでなく、植物も行っており、遺伝子解析の結果[18]によると、植物のセルフィングは百万年前の進化で出現したと推定されている。植物の有性生殖が12億年前に始まったことを考えるなら、セルフィングの歴史は極めて浅いと言わざるを得ない。
進化は一方向的で不可逆的か
高木の「原初有性生殖仮説」は、生物は下等で単純な段階から高等で複雑な段階へと一方向的かつ不可逆的に進化するという考えを前提としている。それゆえ、ゾウリムシにオートガミー/同系交配と異系交配という二つの異なる生殖方法を見つけると、前者から後者が進化したに違いないと決めつけ、さらに下等な段階である無性生殖をオートガミー/同系交配の前に置き、両者を架橋する中間的段階として「原初有性生殖」を想定したのだろう。
高木のそうした偏見は、バクテリアの有性生殖を否定する際にも見られる。高木は、オオアメーバ(Amoeba proteus)のように性分化または雌雄性の存在が認められない真核生物(原生生物)がいることを指摘し、「新規に登場した真核生物で性分化を捨て去るはずがないと考えるのは当然である[19]」と言って、接合するバクテリア(原核生物)の雌雄性に疑問を呈している。真核生物という高等な生物が原核生物という下等な段階から進化したのであるなら、下等な段階で実現した有性生殖が、高等な段階で捨て去るはずがないから、下等な段階で行われていたバクテリアの接合は有性生殖ではないという論理である。
しかし、「新規に登場した真核生物で性分化を捨て去るはずがない」というのはたんなる観念的な思い込みである。ヒルガタワムシ(Bdelloidea)のように、祖先は有性生殖していたにもかかわらず、その後、有性生殖を放棄し、無性生殖しか行わない生物もいる[20]。そのような「後退」は信じられないと言う人は、生命が行っている進化の意義を誤解している。下等で単純な段階から高等で複雑な段階へと進化することは、生物にとって自己目的的に目指すべきものではなく、生き延びるという究極目的のための手段に過ぎないのであって、もしも有性生殖よりも無性生殖の方が有利なら、無性生殖に戻ることは「後退」ではなくて、進化なのである。オートガミーやセルフィングも、典型的な無性生殖から典型的な有性生殖へと進化する過渡期の現象ではなくて、典型的な有性生殖からの進化と見るべきである。
生物が行う進化は、価値的な向上を意味する進歩と区別されるべきである。もしも進化が、価値的に低い状態から高い状態への進歩であるとするなら、なぜ価値的に低い状態の「原始的」な生命が今日に至るまで絶滅することなく生き延びているのかが説明できなくなる。過去の進化の歴史を振り返ればわかるように、むしろ高度に進化した生物の方が、環境の激変に際して絶滅しやすい。それゆえ、下等で単純な段階から高等で複雑な段階へと進化することは、必ずしも進歩とは言えない。もしも生命にとって進歩と呼べるものがあるとするなら、それは進化を通じて、生命全体が多様になることである。多様な種が生まれれば生まれるほど、生命全体が絶滅するリスクが低下するからだ。
オートガミーや同系交配が異系交配の後に始まったとするなら、それにはどのような進化論的な意味があるのか。異系交配による有性生殖は、遺伝子の多様化をもたらすが、有性生殖する生物の数が増えてくると、画一的に多様化することに対するメタレベルの多様化が必要になってくる。オートガミーや同系交配する種が極めて少数である限り、遺伝子を多様化する有性生殖の戦略に大きな影響を与えない。むしろ有性生殖自体の多様化に寄与している。とはいえ、多様化の否定が多様化の肯定になるためには、画一的な多様化がある程度普及してからでなければならない。それが後から派生的に現れた理由であろう。
有性生殖の起源は何か
最後に『有性生殖論』の本来の主題、すなわち、有性生殖の起源は何かという根源的な問いに立ち返ることにしよう。知られている最古の有性生殖生物が紅藻である以上、有性生殖の起源を探る上で参考にしなければならないのは、ゾウリムシではなくて、紅藻である。そして、紅藻には、その点で注目すべき細胞核の細胞間移動という現象が見られる。
以下の図に上から下にかけて経時的に描かれているように、左の寄生紅藻は、右の宿主紅藻に対して、二つある核のうち、一つを宿主紅藻に注入するという、まるでウイルスがやるような遺伝子の水平移動をする。
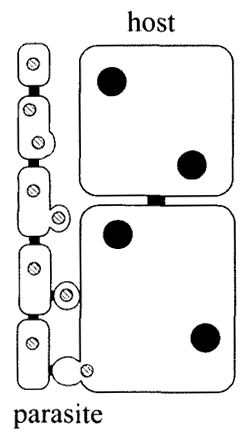
宿主紅藻内に注入された細胞核は、そこで複製され、さらに別の宿主紅藻に注入される。この現象は、バクテリアの接合とよく似ている。例えば、大腸菌は、自分のDNAとは別に、Fプラスミドと呼ばれるDNAを持っていて、Fプラスミドを持っている大腸菌が、持たない大腸菌と接合すると、Fプラスミドが持たない菌に移動する。おそらく、Fプラスミドはもともと大腸菌に感染したウイルスで、ウイルスが伝播するようにFプラスミドは他の大腸菌に伝播しているのだろう。
DNAが移動する大腸菌と細胞核自体が移動する紅藻とでは違うと思うかもしれないが、ワクチニア・ウィルスのように、細胞に侵入後、細胞内の小胞体膜を使って、細胞核のような脂質二重膜で自分を丸く覆い、「ミニ核」ともいうべきものを作るウイルスもいる[21]。寄生紅藻内から移動する細胞核も、こうした類のウイルスが起源と考えれば、その奇妙な振る舞いも理解できる。現在確認されている最古の有性生殖であるウシケノリの生殖は、移動する二つの細胞核がたまたま一つの細胞で一緒になり、二倍体を形成したことで始まったと考えられる。既に見た通り、高木は、トランスポゾン、ウイルス感染、寄生、共生などによる水平遺伝によっても遺伝子情報は多様化するという理由で、遺伝的多様化(ゲノムの多様化)は有性生殖ではないと主張していた。しかし、高木が有性生殖にとって非本質的として切り捨てた水平遺伝にこそ、有性生殖の起源があるのではないのか。
ウィルス(virus)は、もともとラテン語で「毒」を意味する言葉だ。かつてはたんなる病原体としてしか思われていなかったウイルスも、現代の生物学では、生命の進化に大きな役割を果たした準生命体として認識されるようになった。高木は、「突然変異は進化にとっての唯一の原動力[22]」と言うが、この認識はもう古い。生命の起源を説明する最も有力な仮説は、RNAワールド仮説で、それによれば、最初の生命を作ったのは、RNAウイルスである。生命が誕生した後も、ウイルスによる細胞の支配をめぐる攻防は続いた。それはあたかも、最初の王朝が成立した後も、王国の支配をめぐる戦いが続くのと同じである。
王朝は、クーデターや内乱など内部的な出来事で断絶し、新しい王朝に交代することもあれば、異民族による侵略と征服といった外部からの影響で断絶し、国家権力が乗っ取られることもある。内ゲバ型であれ、外ゲバ型であれ、ゲバルト(暴力)が用いられる以上、王朝の終焉は流血の惨事を免れない。これに対して、民主主義国家では、権力者の任期は予め決められており、任期が終われば、平和的な権力移譲が行われ、環境の変化に素早く対応した政策変更が可能である。
国家の支配者と似たことが、細胞の支配者である遺伝子についても言える。無性生殖では、遺伝子は原理的には変化しないはずなのだが、実際は、突然変異によって内的に変わることもあれば、レトロウィルスによる遺伝子の組み込みによって外的に変わることもある。内的な変化であれ、外的な変化であれ、それは暴力的なゲノムの変更であり、生命にとって極めて危険な出来事である。これに対して、有性生殖では、ゲノムの寿命が予め決められており、ゲノムの変更は自律的に行うようにプログラムされている。定期的で自律的なゲノムの変更により、有性生殖する生物は、環境の変化に素早く対応することが可能である。
無性生殖から有性生殖への進化は、同じゲノムを維持し続ける、寿命のない生命の生殖から、ゲノムを多様化する、寿命のある生命の生殖への進化ではなくて、他律的なゲノムの多様化と寿命設定から自律的なゲノムの多様化と寿命設定への進化である。それは、王朝国家から民主主義国家への進化が、権力移譲のない政治システムから権力移譲のある政治システムへの進化ではなくて、事前に決まっていない時期に、暴力的な方法でしか権力者を打倒できない政治システムから、事前に決まっているルールに則って、平和裏に権力移譲が行われる政治システムへの進化であるのと同じことだ。
私は、以前、有性生殖にはパラドキシカルな側面があると書いた。
多くの生物学者(特に利己的遺伝子の仮説を信じている生物学者)は、自分の遺伝子を後世に伝えることが生物の究極の目的だと考えている。そうだとするならば、寄生者対策としての有性生殖は、きわめてパラドキシカルである。ウイルスによって遺伝情報を変えられることを防ぐために、有性生殖によって遺伝情報を変えるということは、暴力団から店を守るために暴力団を用心棒として雇うようなものであるからだ。暴力団に金を渡さないためには、暴力団に金を渡さなければならない。お望みとあらば、雇った暴力団に「警察」あるいは「軍隊」、上納金に「税金」といったもっともらしく聞こえる名前を付けてもよい。[23]
暴力団に金を渡すにしても、予見不可能な時期に暴力的に行われるよりも、予見可能な時期に平和的に行う方が、店にとっては良い。そして、そこに有性生殖の意義がある。
参照情報
- 高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21).
- 高木由臣『生物の寿命と細胞の寿命―ゾウリムシの視点から』平凡社 (1993/9/1).
- 高木由臣『寿命論 細胞から「生命」を考える』NHK出版 (2009/1/27).
- 高木由臣『生老死の進化: 生物の「寿命」はなぜ生まれたか』京都大学学術出版会 (2018/11/9).
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 35.
- ↑Barfooz at the English Wikipedia. “Paramecium aurelia.” Licensed under CC-BY-SA.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 56.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 117.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 93-94.
- ↑高木由臣『寿命論―細胞から「生命」を考える』日本放送出版協会 (2009/01). p. 37.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 91.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 94.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 99.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 101.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 157.
- ↑Johnmartindavies. “Red Algae on bleached coral.” Licensed under CC-BY-SA.
- ↑峯一朗. “紅藻類における受精の形態学的・組織化学的研究." PLANT MORPHOLOGY 9.1 (1997): 43-50. p. 44.
- ↑“Multicellular filaments from the ca. 1200-Ma Hunting Formation (Somerset Island, arctic Canada) are identified as bangiacean red algae on the basis of diagnostic cell-division patterns. As the oldest taxonomically resolved eukaryote on record Bangiomorpha pubescens n. gen. n. sp. provides a key datum point for constraining protistan phylogeny. Combined with an increasingly resolved record of other Proterozoic eukaryotes, these fossils mark the onset of a major protistan radiation near the Mesoproterozoic/Neoproterozoic boundary. Differential spore/gamete formation shows Bangiomorpha pubescens to have been sexually reproducing, the oldest reported occurrence in the fossil record.” ― Butterfield, Nicholas J. “Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes." Paleobiology 26.3 (2000): 386-404.
- ↑“Detailed data on bacterial and protistan microfossils are presented from a 0.003 mm3 piece of Triassic amber (Schlierseerit, Upper Triassic period, 220-230 million years old).” Schönborn, Wilfried, Heinrich Dörfelt, Wilhelm Foissner, Lothar Krienitz, and Ursula Schäfer. “A Fossilized Microcenosis In Triassic Amber.” Journal of Eukaryotic Microbiology 46, no. 6 (November 1, 1999): 571–84.
- ↑“Ciliates, a major eukaryotic crown-group lineage with thousands of living species, are poorly represented in the fossil record. Ciliate biomarkers are known from the Precambrian, but only one group, the tintinnids, have an extensive fossil record dating back to the Ordovician. Thus, the occurrence of probable ciliate body fossils in Neoproterozoic rocks confirms their earlier appearance, so far inferred only from molecular sequence data and biomarkers. In this paper, we describe those fossils from the 580 million year old Precambrian Doushantuo phosphates, Guizhou, South China.” ― Li, C.-W.; et al. (2007). “Ciliated protozoans from the Precambrian Doushantuo Formation, Wengan, South China“. Geological Society, London, Special Publications. 286: 151–156.
- ↑Droser, Mary L., and James G. Gehling. “Synchronous aggregate growth in an abundant new Ediacaran tubular organism." Science 319.5870 (2008): 1660-1662.
- ↑“An analysis of the genome-wide pattern of linkage disequilibrium suggests that selfing most likely evolved roughly a million years ago or more.” ― Tang, Chunlao, et al. “The Evolution of Selfing in Arabidopsis thaliana." Science 317.5841 (2007): 1070-1072.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 91.
- ↑“We conducted a molecular genetic test of this indication that bdelloid rotifers may have evolved without sexual reproduction or genetic exchange. The test is based on the expectation that after millions of years without these processes, genomes will no longer contain pairs of closely similar haplotypes and instead will contain highly divergent descendants of formerly allelic nucleotide sequences. We find that genomes of individual bdelloid rotifers, representing four different species, appear to lack pairs of closely similar sequences and contain representatives of two ancient lineages that began to diverge before the bdelloid radiation many millions of years ago when sexual reproduction and genetic exchange may have ceased.” ― Welch, David B. Mark, and Matthew Meselson. “Evidence for the Evolution of Bdelloid Rotifers Without Sexual Reproduction or Genetic Exchange." Science 288.5469 (2000): 1211-1215. ヒルガタワムシは輪形動物門に属し、ヒルガタワムシ鋼以外は有性生殖する。
- ↑“Vaccinia virus (vv), a member of the poxvirus family, is unique among most DNA viruses in that its replication occurs in the cytoplasm of the infected host cell. Although this viral process is known to occur in distinct cytoplasmic sites, little is known about its organization and in particular its relation with cellular membranes. The present study shows by electron microscopy (EM) that soon after initial vv DNA synthesis at 2 h postinfection, the sites become entirely surrounded by membranes of the endoplasmic reticulum (ER).” ― Tolonen, Nina, et al. “Vaccinia Virus DNA Replication Occurs in Endoplasmic Reticulum-enclosed Cytoplasmic Mini-Nuclei." Molecular biology of the cell 12.7 (2001): 2031-2046.
- ↑高木由臣『有性生殖論―「性」と「死」はなぜ生まれたのか』日本放送出版協会 (2014/1/21). p. 138.
- ↑永井俊哉. “有性生殖はなぜ必要なのか.” 2004年10月16日.
{kind=link}
{kind=link}